4.1. Aspectos morfológicos.
4.1.1. La cara y los dientes.
Un hecho característico del proceso de hominización es la disminución gradual del tamaño de la cara y de los dientes. Todos los grandes simios están dotados de enormes caninos (colmillos) que destacan del resto de los dientes. A medida que avanzamos en el proceso de hominización, se observa que los caninos van reduciéndose de tamaño. Además, los dientes que sirven para masticar (premolares y molares) han ido disminuyendo su tamaño progresivamente. Estos cambios provocan una disminución del tamaño de la cara y de las mandíbulas. La cara de los primeros antepasados del ser humano era grande y estaba situada al frente del cráneo. A medida que los dientes se redujeron y el cerebro aumentó, la cara disminuyó y varió su posición; así, la cara de los seres humanos modernos está situada debajo, no delante, del cerebro. Otros rasgos: en el ser humano actual destacan la nariz y el mentón en el perfil de la cara.
4.1.2. Liberación de los miembros superiores.
La postura bípeda dejó libres los miembros superiores que ya no tienen que cumplir la función de patas (excepto en los niños muy pequeños) ni la de braquiación, es decir, el desplazamiento de rama en rama con los brazos, aun cuando la actual especie humana de la cintura hacia arriba mantenga una complexión de tipo arborícola.
Esta liberación de los miembros superiores fue, en su inicio, una adaptación óptima al bioma de sabana; al marchar bípedamente y con los brazos libres, los ancestros del hombre podían recoger más fácilmente su comida: raíces, frutos, hojas, insectos, huevos, reptiles pequeños, roedores y carroña. En efecto, muchos indicios hacen suponer como probable que nuestros ancestros fueran en gran medida carroñeros y, dentro del carroñeo, practicaran la modalidad llamada cleptoparasitismo, esto es, robaban las presas recién cazadas por especies netamente carnívoras; para tal práctica, nuestros ancestros debían haber actuado en bandas, organizadamente.
Los miembros superiores, siempre en relación con otras especies, se han acortado. Estos miembros superiores, al quedar liberados de funciones locomotoras, se han podido especializar en funciones netamente humanas. El pulgar oponible es una característica heredada de los primates más antiguos, pero si en éstos la función principal ha sido la de aferrarse a las ramas y en segundo lugar aprehender las frutas o insectos que servían de alimento, en la línea evolutiva que desemboca en nuestra especie la motilidad de la mano, y en particular de los dedos de ésta, se ha hecho gradualmente más precisa y delicada, lo que ha facilitado la elaboración de artefactos. Aún no se tiene conocimiento respecto al momento en que la línea evolutiva comenzó a crear artefactos, aunque es seguro que ya hace más de 2 millones de años Homo habilis/Homo rudolfensis realizaba toscos instrumentos que utilizaba asiduamente (en todo caso, los chimpancés, en estado silvestre, confeccionan "herramientas" de piedra, madera y hueso muy rudimentarias). El desarrollo de la capacidad de pronación en la articulación de la muñeca también ha sido importantísimo para la capacidad de elaborar artefactos.
 El origen de la mano humana.
El origen de la mano humana.Por David M. Alba, Salvador Moyá y Meike Köhler (febrero de 2005) [Saltarse este artículo]
Una de las principales críticas a las que tuvo que hacer frente Darwin tras la publicación de El Origen de las Especies sostenía que la selección natural sólo podía eliminar los caracteres inadecuados, pero no crear las complejas adaptaciones que observamos en la naturaleza. Según esta crítica los estadios primeros, en sí mismos incompletos, no podían conferir la ventaja adaptativa que proporciona la estructura acabada. En ediciones posteriores del libro, Darwin rechazó la objeción apelando al principio de cooptación.
De acuerdo con el principio de cambio funcional, como se conoce también el de cooptación, en sus estadios iniciales la estructura habría desempeñado una función distinta; ahora bien, tal función del pasado habría sido, tiempo después, "cooptada", vale decir, reclutada para cumplir la misión actual. Para esa desconexión entre el origen de una estructura y su función actual, los paleontólogos Stephen Jay Gould y Elisabeth Vrba acuñaron en 1982 el término exaptación. Debía aplicarse a aquellas estructuras cooptadas, es decir, aquellas que en su origen o no desarrollaban ninguna función o desarrollaban una función adaptativa diferente de la que, andando el tiempo, cumplirían. Había, pues, que distinguir entre adaptaciones genuinas, propias de las estructuras que evolucionaron mediante selección natural para llevar a cabo su función actual, y exaptaciones.
Adaptación y exaptación
No resulta fácil saber cuándo nos encontramos ante una adaptación y cuándo ante una exaptación. En paleontología no podemos observar de un modo directo la función de una estructura, sino que hemos de inferirla a partir de la forma. Si interpretamos todas las estructuras como adaptaciones, podremos deducir la función de las estructuras fósiles a partir de la función que desempeñan en los descendientes. Pero si hemos de separar, de acuerdo con el principio de cooptación, función actual y origen evolutivo, nos encontramos con un problema importante: ¿cómo averiguar la función de una estructura en el registro fósil?
Para aceptar una hipótesis de adaptación en el registro fósil, señala George Lauder, se requiere, como mínimo, que exista una concordancia histórica entre el origen de la estructura y el inicio de una presión selectiva que favorezca la función en cuestión. En palabras de E. N. Arnold: "Queda descartada cualquier hipótesis de adaptación si aparece el carácter novedoso antes que el pertinente régimen selectivo". A menudo se confunde exaptación con adaptación, pese a la claridad del principio: si la aparición de la nueva forma y de la nueva función no coinciden, nos hallamos ante una exaptación.
Enunciadas tales premisas conceptuales, veamos su aplicación al origen de la mano humana, uno de nuestros objetos de estudio en el Instituto de Paleontología M. Crusafont de Sabadell. ¿Evolucionaron las proporciones de la mano humana en el sentido de una adaptación para la fabricación de herramientas? ¿O ya existían anteriormente para desarrollar una función distinta? Dentro del orden de los Primates, los humanos modernos (Homo sapiens) presentan dos características distintivas principales, a saber, la locomoción bípeda y un grado elevado de encefalización. Ambas peculiaridades aparecieron en algún punto de la evolución humana después de que el linaje humano se separase del linaje de los grandes antropomorfos. El bipedismo se remonta como mínimo hasta nuestros ancestros australopitecos (género Australopithecus); la encefalización incrementada, a los primeros representantes del género Homo.
La aparición del bipedismo permitió que las extremidades anteriores, en particular las manos, quedaran liberadas de funciones locomotoras y se aplicaran exclusivamente a la manipulación. A su vez, el elevado grado de capacidad de procesar información, asociado a un gran tamaño del encéfalo en relación con la masa corporal, posibilitó, en el curso de la evolución humana, no sólo usar, sino también fabricar, útiles cada vez más complejos, hasta desembocar en las muy tecnificadas sociedades actuales. Pero ello no se habría logrado sin una morfología adecuada de la mano humana, que permitiese una manipulación eficiente durante el uso y la fabricación de instrumentos líticos.
Pinzas de precisión
Entre las diversas características de la mano humana que facilitan la manipulación de útiles, destacan las proporciones; en particular, la gran longitud del pulgar con respecto a la longitud total de la mano. Tales proporciones son esenciales para realizar la "pinza de precisión de tipo humano". Los primates en general pueden llevar a cabo dos tipos de pinza: de precisión y de fuerza. Mediante la primera, sujetan los objetos entre el pulgar y uno o más del resto de dedos de la mano. Mediante la pinza de fuerza, el individuo sujeta los objetos entre la palma de la mano y el resto de dedos, con el pulgar sirviendo de mero soporte.
Humanos y grandes antropomorfos (chimpancés, bonobos, gorilas y orangutanes) realizan ambos tipos de operaciones. Sus manos utilizan, pues, pinzas de precisión y de fuerza en distintas proporciones. Sólo los humanos, sin embargo, realizan un tipo particular de pinza de precisión, la "pinza de precisión yema con yema". Aquí, el objeto en cuestión se sujeta con fuerza entre las yemas del pulgar y de uno o más dedos de la mano (normalmente el índice). Este tipo de pinza, el más habitual entre los humanos, resulta vedado a los grandes antropomorfos por una razón biomecánica: la escasa longitud del pulgar con respecto al resto de los dedos.Las manos de los grandes antropomorfos, largas en comparación con las de los humanos, resultan ser una adaptación a la locomoción arborícola. Sirven para suspenderse de las ramas y trepar por los troncos. Tanto el orangután como los simios antropomorfos africanos presentan este patrón de manos largas con un pulgar corto. Por tanto, con los elementos de juicio disponibles, podemos suponer que la mano corta y el pulgar largo de los humanos constituyen una característica derivada de nuestro linaje, evolucionada a partir de un antepasado con proporciones de la mano más simiescas, más primitivas.
Las proporciones de la mano de los simios antropomorfos no les impiden utilizar, e incluso fabricar, instrumentos, lo mismo en libertad que en cautividad; existen pruebas abundantes del uso de instrumentos por diversas especies de primates. En el caso particular de los chimpancés, se conoce incluso cierta variabilidad de un grupo a otro; además, son los únicos que no se limitan a aprovechar los objetos naturales encontrados, sino que los modifican. Mas, para esa fabricación de instrumentos toman tan sólo materiales orgánicos (ramas, por ejemplo), que no suelen dejar huella en el registro arqueológico.
Fabricación de útiles
La fabricación de instrumentos líticos y el uso de instrumentos secundarios (útiles para hacer útiles) corresponden de forma exclusiva a los humanos. El paleoantropólogo Randall L. Susman ha asociado las características morfológicas que permiten llevar a cabo una pinza de precisión de tipo humano con una mayor capacidad de manipular instrumentos. Más aún, erige esas características en "criterio de fabricación de instrumentos".
Si Susman tuviera razón, las proporciones de la mano humana deberían ser una adaptación para la fabricación de instrumentos líticos, cuando menos. La hipótesis entra, desde luego, dentro de lo posible. Pero el principio de cooptación nos advierte de un error potencial: igualar la función actual de una estructura anatómica con su origen evolutivo. ¿Son las proporciones de la mano humana una adaptación para la fabricación de instrumentos líticos? Para responder a esta pregunta, se requiere verificar el criterio de concordancia histórica, esto es, averiguar en qué momento evolucionaron las proporciones de la mano humana y en qué momento se originaron las presiones de selección impuestas por la fabricación de instrumentos líticos. Sólo el registro fósil puede suministrarnos tales datos.
Lo que dice el registro fósil
El registro fósil del comportamiento humano, o registro arqueológico, indica que las industrias líticas más antiguas documentadas datan de hace 2,5 millones de años. Por desgracia, a causa de la escasez de huesos de la mano de humanos fósiles, no se ha podido documentar todavía la transición evolutiva desde una mano simiesca hasta una mano humana. Por un lado, existen fósiles de la mano de Ardipithecus ramidus, uno de los miembros más antiguos reconocidos del linaje humano, hallado en Etiopía, pero aún no han sido publicados. Se cuenta también con una mano articulada, asociada a un esqueleto de Australopithecus africanus, recuperado en la cueva de Sterkfontein, aunque hasta la fecha sólo se han publicado informes preliminares. El espécimen manual más completo del registro fósil humano, el llamado O.H.7, procedente de la garganta de Olduvai, no presenta ni el metacarpiano ni la falange proximal del pulgar; no aporta, pues, suficiente información con que reconstruir las proporciones de la mano.
Quedan, por fin, los restos de la mano de Australopithecus afarensis (la especie del famoso esqueleto parcial de "Lucy"), desenterrados en el yacimiento etíope de A.L.333/333w. Entrañan un grave problema: los metacarpianos y falanges corresponden a varios individuos mezclados. Tal desorden ha dado pie a una doble interpretación de las afinidades morfológicas de la mano de Australopithecus. Para ciertos autores, incluido Susman, se trataría de una mano esencialmente simiesca y adaptada a la suspensión arborícola; además, al ser una mezcolanza de huesos de distintos individuos, no podrían determinarse las proporciones del pulgar. Para Mary W. Marzke y otros, la mano de Australopithecus afarensis mostraría algunos caracteres más cercanos a los humanos; el pulgar en particular sería más largo que en los grandes simios antropomorfos, sin alcanzar el tamaño de los humanos modernos y, por tanto, excesivamente corto para permitir la pinza de precisión yema con yema.
En cualquier caso, la mezcla de huesos constituía en sí misma un problema. Los partidarios de la hipótesis simiesca podían argumentar que las proporciones más humanas esgrimidas por los partidarios de la segunda hipótesis resultaban de la combinación aleatoria de los huesos.
Aleatorización
Los autores de este artículo resolvimos la objeción de la mezcla de huesos en un artículo reciente, aparecido en Journal of Human Evolution. Basamos nuestro estudio de las proporciones de la mano de Australopithecus afarensis en la reconstrucción de dos manos parciales (izquierda y derecha) atribuidas a un mismo individuo. Cuando los huesos de la mano derecha e izquierda se combinan, a la reconstrucción sólo le falta la falange media del dedo cuarto; ello nos permite comparar las proporciones manuales de Australopithecus afarensis con humanos modernos y grandes antropomorfos actuales.
El supuesto de que los huesos seleccionados pertenecen a un mismo individuo, y los descartados no, se fundamentó en la compatibilidad de tamaños y en la estrecha congruencia encontrada entre las facetas articulares. De ello podía extraerse una hipótesis verosímil de trabajo, pero no una demostración irrefutable. En efecto, la mayoría de los fósiles (además de los huesos de la mano hay otros restos óseos) no se encontraron en conexión anatómica; se hallaban dispersos por una ladera de pocos metros cuadrados. Hay representados restos de, al menos, tres individuos de tamaño grande y dos de tamaño pequeño.
Nuestra principal aportación consistió en haber demostrado estadísticamente (con una fiabilidad del 95%) que las conclusiones obtenidas suponiendo que los huesos empleados en la reconstrucción pertenecían a un mismo individuo seguirían siendo ciertas, aun cuando estos huesos perteneciesen todos a individuos distintos. Esta demostración estadística, clave para dar suficiente robustez a nuestras conclusiones, se llevó a cabo mediante la aplicación de un proceso de aleatorización a las técnicas estadísticas estándar empleadas.
En esencia, se acometieron los análisis no sólo con la mano compuesta de Australopithecus afarensis, sino también con una serie de manos "quiméricas" de humanos y grandes antropomorfos actuales. Las quimeras se generaron de una forma aleatoria, mediante la mezcla de huesos procedentes de muchos individuos distintos. A través de ese método pudo calcularse la probabilidad de obtener erróneamente las proporciones de Australopithecus afarensis al mezclar aleatoriamente huesos correspondientes a individuos distintos. Cuando esta probabilidad no llegó al 5%, quedó estadísticamente descartada la posibilidad de que los resultados obtenidos se debieran al azar.
Se comparó la mano de Australopithecus afarensis con manos de hominoideos actuales (humanos, chimpancés, bonobos, gorilas y orangutanes). En ese cotejo se tuvieron en cuenta la longitud relativa del pulgar y las proporciones globales de la mano. En el proceso de comparación se emplearon residuos alométricos, que permiten calcular la desviación de un individuo con respecto a la tendencia general de la muestra en cuanto al tamaño corporal; se sometieron los datos a un análisis de la varianza y análisis multivariante canónico, también llamado análisis discriminante.
En lo concerniente a las proporciones globales de la mano, Australopithecus afarensis se situó más cerca de los humanos que de los simios antropomorfos, aunque no sin diferencias estadísticamente significativas con los humanos actuales, una discrepancia explicable al tratarse de géneros distintos. El análisis de aleatorización confirmó tales resultados. Mostró, en particular, que, si se tiene en cuenta el grado de similitud con los humanos actuales, puede excluirse que las proporciones de tipo humano de la mano de Australopithecus afarensis se deban a la mezcla de huesos procedentes de distintos individuos. En lo concerniente al tamaño relativo del pulgar, Australopithecus afarensis también se aproximaba bastante a los humanos con un pulgar muy alargado, en comparación con los simios antropomorfos. También lo confirmó el procedimiento de aleatorización.
Atendiendo a la masa corporal, el largo pulgar de los humanos no resulta tanto de un alargamiento en términos relativos cuanto de un acortamiento del resto de la mano; idéntica explicación cabe ofrecer del pulgar de Australopithecus afarensis. Tales afirmaciones han de tomarse con cautela, dada la falta de conocimientos seguros sobre el tamaño corporal real de los fósiles; con todo, el análisis de aleatorización revela que sí podemos llegar, al menos, a conclusiones firmes respecto el acortamiento de la mano.
El hecho de que Australopithecus afarensis preceda en alrededor de un millón de años los primeros restos arqueológicos de industria lítica, fechados hace 2,5 millones de años, nos permite refutar la hipótesis de Susman. Según éste, las proporciones humanas de la mano evolucionaron en origen como una adaptación al uso y la fabricación de instrumentos líticos.
En efecto, queda patente que no se cumple el criterio de concordancia histórica entre el origen del carácter (las proporciones de la mano humana) y el comienzo de la presión selectiva pertinente (la fabricación de instrumentos líticos). Ello no excluye que se seleccionaran otras características de la mano humana para cumplir dicha función. Ahora bien, en lo concerniente a las proporciones de la mano, la fabricación de instrumentos líticos habría favorecido como máximo pequeños reajustes de las proporciones en cuestión. Con otras palabras, las proporciones peculiares de la mano humana constituirían una adquisición anterior a la fabricación de útiles de piedra, adquisición que habría sido cooptada luego para llevar a cabo la nueva función. Con respecto a la fabricación de instrumentos líticos, las proporciones de la mano humana no serían una adaptación, sino una exaptación.
Origen de las proporciones de la mano humana
Una vez descartada la fabricación de instrumentos líticos, falta por resolver el origen de las proporciones humanas de la mano. De acuerdo con cierta hipótesis, que no puede comprobarse en el registro arqueológico, debería atribuirse a las presiones de selección impuestas por tipos de manipulación más complejos que los observados entre primates no humanos actuales. Razona la hipótesis que los homininos primitivos (los antecesores inmediatos de los humanos) habrían mostrado comportamientos instrumentales más complejos que los actuales chimpancés en libertad, incluyendo el uso y tal vez fabricación de útiles con materiales orgánicos. Esa argumentación introduce un escenario posible, aunque no necesario. Atendamos a los primates no humanos actuales. Las manos les sirven para un amplio abanico de diversas funciones locomotoras y manipuladoras. Desde nuestra perspectiva antropocéntrica se nos hace a veces difícil entender que, en los precursores del linaje humano, la manipulación pudiera estar desvinculada de la fabricación de instrumentos. No obstante, todos los primates utilizan sus manos para tareas muy dispares, desde el acicalamiento hasta la recolección y el procesado de alimentos. Esas operaciones, sin relación con las herramientas, requieren, sin embargo, movimientos complejos.
Si se quiere evitar caer en un razonamiento circular, hay que abordar el origen de la mano humana en un marco que trascienda la vinculación de la misma con el manejo de útiles. En ese nuevo enfoque, deberían considerarse otras actividades llevadas a cabo por los primates en contextos no locomotores y que implican una manipulación compleja.
Aunque los biólogos se empeñen en analizar por separado la evolución de cada carácter, no debe olvidarse que los organismos presentan un alto nivel de integración. Merced a ello, una misma estructura anatómica desempeña a menudo más de una función; en razón de ello también, presiones selectivas de distinta índole actúan a la vez sobre la misma estructura anatómica. Lo observamos en las manos: aunque manipulación y locomoción no sean funciones excluyentes, acostumbran implicar presiones selectivas distintas, si no opuestas.
Las presiones selectivas de tipo locomotor ejercen, en general, una influencia más determinante y se reflejan directamente en la morfología de la mano. Las adaptaciones asociadas a la manipulación vienen condicionadas por las restricciones biomecánicas impuestas por la locomoción; con otras palabras: la locomoción podría impedir la adquisición de proporciones óptimas para una manipulación más eficiente. Los primates tienen las manos adecuadas para su tipo de locomoción. Dada la primacía de esa función, acometen como pueden la otra función, la de manipulación.
En los monos cuadrúpedos, las proporciones de la mano son en general más humanas (y por lo tanto, más eficientes para la manipulación), pues el cuadrupedismo terrestre favorece unas manos cortas. Pero en los gibones y orangutanes, con comportamientos locomotores arbóreos de tipo suspensor y trepador, se favorece una mano larga, menos adecuada para la manipulación; lo mismo se aplica a los antropomorfos africanos, que presentan un característico "andar de nudillos de tipo cuadrúpedo".
Sólo la adquisición de un bipedismo terrestre habitual, como encontramos en los humanos modernos, libera las manos de las fatigas locomotoras; quedan exclusivamente expuestas a la presión de selección relacionada con la función manipuladora.
Nuestra hipótesis
Desde los tiempos de Darwin viene admitiéndose una relación entre el bipedismo y la aparición de las aptitudes manipuladoras humanas. Nosotros proponemos la hipótesis de que las proporciones de la mano humana, incluida una longitud notable del pulgar, podrían haberse originado de las mismas presiones de selección que se pueden encontrar entre los primates no humanos actuales, una vez que las manos quedaron liberadas de demandas locomotoras al adoptar la locomoción bípeda.
La hipótesis se revela congruente con los datos anatómicos recabados de Australopithecus afarensis. Además de proporciones manuales de tipo humano, este australopiteco muestra ya muchas adaptaciones a un bipedismo habitual. Algunas de tales adaptaciones reflejan una menor eficacia para trepar a los árboles. Nuestra hipótesis concuerda también con otro aspecto de interés en Australopithecus afarensis: el incremento relativo de la longitud del pulgar no se obtuvo alargando el pulgar, sino reduciendo la longitud de la mano, que es lo que cabría esperar de una mano no sometida a una intensa presión de selección de tipo locomotor.
A propósito del tipo de cuadrupedismo terrestre en los antropomorfos africanos, podría aducirse que se privilegió un andar de nudillos frente al bipedismo o el cuadrupedismo digitígrado por una razón de peso: para conservar una mano larga, exigida en el mantenimiento de comportamientos arborícolas ancestrales, pero poco adecuada para una manipulación eficiente. Muchas de las características bípedas de Australopithecus afarensis implican ya un decremento en la eficiencia de la locomoción arbórea. Ello se hace evidente en los brazos, que tienen escasa incidencia en la mecánica del bipedismo, pero desempeñan una función primordial en la locomoción arbórea.
Nuestra explicación encaja también con el descubrimiento de que Oreopithecus bambolii, un simio antropomorfo bípedo, que evolucionó en condiciones de insularidad en el Mioceno Superior de Italia, tenía un pulgar de proporciones humanas. Una vez más, la liberación de los requerimientos locomotores facilita la aparición de unas proporciones manuales de tipo humano como resultado de presiones de selección relacionadas con la manipulación, pero desligadas de la fabricación de instrumentos.4.1.3. Visión.
El humano hereda de los prosimios la visión estereoscópica y pancromática (la capacidad de ver una amplia tonalidad de los colores del espectro visible). Los ojos en la parte delantera de la cabeza posibilitan la visión estereoscópica (en tres dimensiones), pero si esa característica surge en los prosimios como una adaptación para moverse mejor en el medio arbóreo, en Homo sapiens tal función cobra otro valor: facilita la mirada a lontananza, el otear horizontes. En este aspecto la visión es bastante más aguda en los humanos que en los otros primates y en los prosimios. Esto facilitará el hecho por el cual Homo sapiens sea un ser altamente visual (por ejemplo las comunicaciones mediante la mímica), facilitará asimismo lo imaginario.
4.1.4. Especialización.
Pese al conjunto de modificaciones morfológicas antes reseñadas, desde el punto de vista de la anatomía comparada, llama la atención una cuestión: Homo sapiens es un animal relativamente poco especializado. En efecto, gran parte de las especies animales ha logrado algún tipo de especialización anatómica (por ejemplo los artiodáctilos poseen pezuñas que les permiten correr en las llanuras despejadas), pero las especializaciones, si suelen ser una óptima adaptación a un determinado bioma, conllevan el riesgo de la desaparición de la especie especializada y asociada a tal bioma si éste se modifica.
La ausencia de tales especializaciones anatómicas ha facilitado a los humanos una adaptabilidad inusitada entre las demás especies de vertebrados para adecuarse a muy diversas condiciones ambientales.
Más aún, aunque parezca paradójico, Homo sapiens tiene características neoténicas. En efecto, la estructura craneal de un Homo sapiens adulto se aproxima más a la de la cría de un chimpancé que a la de un chimpancé adulto: el rostro es achatado (ortognato o de bajo índice facial) y es casi inexistente el torus supraorbitario (en la humanidad actual apenas se encuentran vestigios de torus en las poblaciones llamadas australoides). De otro modo se puede decir que los arcos superciliares de Homo sapiens son infantiles, delicados, el rostro aplanado o ligeramente prognato.
Asociado al hecho por el cual morfológicamente el ser humano tenga características que le aproximan a las de un chimpancé inmaduro se encuentra el ortognatismo y esto quiere decir, entre otras cuestiones, que los dientes de Homo sapiens son relativamente pequeños y poco especializados; las mandíbulas, por esto, se ha abreviado y hecho más delicadas. Falta además el diastema o espacio en donde encajan los colmillos. La debilidad de las mandíbulas humanas las hace casi totalmente inútiles para la defensa a mordiscos ante un predador y, asimismo, son muy deficientes para poder consumir gran parte del alimento en su estado natural, lo que es uno de los muchos déficits corporales que llevan al humano a vivir en una sociedad organizada.
4.2. Bipedestación.
Los homininos, primates bípedos, habrían surgido hace unos 6 ó 7 millones de años en África, cuando dicho continente se encontró afectado por una progresiva desecación que redujo las áreas de bosques y selvas. Como adaptación al bioma de sabana aparecieron primates capaces de caminar fácilmente de modo bípedo y mantenerse erguidos. Más aún, en un medio cálido y con fuerte radiación ultravioleta e infrarroja, una de las mejores soluciones adaptativas son la marcha bípeda y la progresiva reducción de la capa pilosa, lo que evita el excesivo recalentamiento del cuerpo. Hace 150.000 años el norte de África volvió a sufrir una intensa desertización, lo cual significó otra gran presión evolutiva como para que se fijaran los rasgos principales de la especie Homo sapiens.
Para lograr la postura y marcha erecta han tenido que aparecer importantes modificaciones:
- Cráneo. Para permitir la bipedestación, el foramen magnum se ha desplazado; mientras en los simios el foramen magnum se ubica en la parte posterior del cráneo, en el Homo sapiens (y en sus ancestros directos) el foramen magnum se ha desplazado casi hacia la base del mismo.
- Columna vertebral. La columna vertebral, bastante rectilínea en los simios, ha adquirido en el Homo sapiens y en sus ancestros bípedos diversas curvaturas que, gracias a un efecto resorte, permiten soportar mejor el peso de la parte superior del cuerpo. Por lo demás la columna vertebral ha podido erguirse casi 90º a la altura de la pelvis. Si se compara con un chimpancé se nota que, al carecer este primate de la curva lumbar, su cuerpo resulta empujado hacía adelante por el propio peso; en la raquis humana el centro de gravedad se ha desplazado, de modo que el centro de gravedad de todo el cuerpo se sitúa encima del soporte que constituyen los pies. Al tener el Homo sapiens una cabeza relativamente grande el centro de gravedad corporal es bastante inestable (y hace que al intentar nadar, el humano tienda a hundirse "de cabeza"). Otro detalle es que las vértebras humanas son más circulares que las de los simios, lo que les permite soportar mejor el peso vertical.

- Pelvis. La pelvis se ha debido ensanchar, lo cual ha sido fundamental en la evolución de nuestra especie. Los huesos ilíacos de la región pelviana en los Homo sapiens (e inmediatos antecesores) "giran" hacia el interior de la pelvis, lo que le permite soportar mejor el peso de los órganos al estar en posición erecta. La citada modificación de la pelvis implica una disminución importante en la velocidad posible de la carrera por parte de los humanos. La bipedestación implica una posición de la pelvis que hace que las crías nazcan "prematuras": en efecto, el parto humano es denominado ventral acodado ya que existe casi un ángulo recto entre la cavidad abdominal y la vagina, que en el pubis de la mujer es casi frontal. Mientras que en todos los otros mamíferos el llamado canal de parto es muy breve, en las hembras de Homo sapiens es muy prolongado y sinuoso, lo que hace dificultosos los alumbramientos. Como se verá más adelante, esto ha sido fundamental en la evolución de nuestra especie.
- Piernas. También para la bipedestación ha habido otros cambios morfológicos muy importantes y evidentes, particularmente en los miembros y articulaciones. Los miembros inferiores se han robustecido, el fémur humano se inclina hacia adentro, de modo que le posibilita la marcha sin necesidad de girar casi todo el cuerpo y la articulación de la rodilla se ha vuelto casi omnidireccional. Aunque en los simios existe una mayor flexibilidad de la articulación de la rodilla, es para un mejor desplazamiento por las copas de los árboles, es así que el humano a diferencia de sus parientes más próximos no marcha con las rodillas dobladas.
- Pies. En los humanos los pies se han alargado, particularmente en el talón, reduciéndose algo los dedos del pie y dejando de ser oponible el pulgar del pie. En líneas generales el pie ha perdido casi totalmente la capacidad de aprehensión. Se sabe, en efecto, que el pie humano ha dejado de estar capacitado para aferrarse a las ramas, pasando en cambio a tener una función importante en el soporte de todo el cuerpo. El dedo mayor del pie tiene una función vital para lograr el equilibrio de los homininos durante la marcha y la postura erecta; en efecto, el pulgar del pie de un chimpancé es transversal, lo que permite a este homínido aferrarse más fácilmente a las ramas, en cambio el pulgar del pie humano, al estar alineado, facilita el equilibrio y el impulso hacia adelante al marchar o correr. Los huesos de los miembros inferiores son relativamente rectilíneos en comparación con los de otros primates.
4.2.1. Origen de la bipedestación.
Existen diferentes hipótesis sobre el por qué los australopitecinos se separaron de los simios iniciando así el curso de la evolución humana. Prácticamente todas las hipótesis sugieren que el cambio medioambiental fue un factor importante, especialmente al favorecer la evolución de la bipedestación. Entre las hipótesis más coherentes se encuentran la de la sabana, la del mosaico de bosques y la de la variabilidad.
Cerca del final del Mioceno, hace entre 8 y 5 millones de años, el clima de la Tierra sufrió un intenso enfriamiento y se hizo más seco. Según la hipótesis de las sabanas, al comenzar este cambio climático se fueron reduciendo las áreas de bosques africanos y a medida que los bosques fueron disminuyendo, una población de simios de África Oriental se fue quedando aislada de las demás poblaciones de simios que vivían en las áreas de mayor densidad de bosques del oeste de África. La población de África Oriental tuvo que adaptarse a un entorno más seco, con mayores áreas de sabanas. Esta hipótesis, como ya se ha dicho, fue conocida como East Side Story.
El aumento de los terrenos secos favoreció la evolución de la vida sobre el suelo e hizo que cada vez fuera más difícil sobrevivir en los árboles. Los simios terrestres pudieron haber formado grandes grupos sociales para favorecer su capacidad de encontrar y recolectar alimentos y defenderse de los predadores, actividades que también pueden haber requerido el desarrollo de una buena comunicación. Las dificultades de la vida en la sabana pudieron asimismo haber fomentado el inicio del uso de utensilios para fines tales como la extracción de carne de las presas.
Estos importantes cambios evolutivos tuvieron que depender de un aumento de la capacidad mental y, por lo tanto, pueden haber estado relacionados con el desarrollo de un cerebro más grande.
Las críticas a la hipótesis de las sabanas son variadas pero se basan principalmente en dos razones: primero, el descubrimiento en 1994 de fósiles de australopitecinos en Chad, África Central, por un equipo científico sugiere que los entornos de África Oriental tal vez no estuvieran totalmente aislados de los que se encontraban más al oeste; segundo, investigaciones recientes sugieren que las sabanas abiertas no aparecieron de forma significativa en África hasta casi 2 millones de años después. Las críticas a la teoría de las sabanas han hecho surgir un gran número de hipótesis alternativas sobre los orígenes de la evolución humana.
La hipótesis del mosaico de bosques sostiene que los primeros australopitecinos evolucionaron en áreas boscosas que formaban parte de un mosaico de bosques y sabanas que les permitían alimentarse tanto en el suelo como en los árboles, y que la alimentación en el suelo favoreció la bipedestación.
La hipótesis de la variabilidad sugiere que, debido a los numerosos cambios en su entorno, los primeros australopitecinos acabaron por vivir en diferentes hábitats, incluidos selvas, bosques abiertos y sabanas. Como consecuencia, sus poblaciones tuvieron que adaptarse a entornos diferentes. Los científicos han demostrado que esta serie de hábitats existían en el momento en que comenzó la evolución de los primeros australopitecinos. De esta forma, el desarrollo de nuevas características anatómicas (en particular la bipedestación), combinadas con la capacidad de trepar a los árboles, pueden haber conferido a los homínidos la versatilidad necesaria para vivir en hábitats diferentes.
4.2.2. Ventajas de la bipedestación.
Es evidente que la gran cantidad de modificaciones anatómicas que condujeron del cuadrupedismo al bipedismo requirió una fuerte presión selectiva. Se ha discutido mucho sobre la ineficacia de la marcha bípeda comparada con la cuadrúpeda. También se ha criticado que ningún otro animal de los que se adaptaron a la sabana al final de Mioceno desarrolló una marcha bípeda. Hemos de tener en cuenta que partimos de homínidos con un tipo de desplazamiento cuadrúpedo que no es ninguna maravilla adaptativa; en efecto, el modo en que se desplazan los chimpancés, apoyando la segunda falange de los dedos de las manos, no puede compararse a la marcha cuadrúpeda de ningún otro mamífero. Puede ser suficiente en trayectos cortos por el suelo, pero no es muy eficaz para grandes desplazamientos en terreno abierto. Los primeros homínidos de sabana probablemente se vieron obligados a desplazarse distancias considerables en campo abierto para alcanzar grupos de árboles situados a distancia. La marcha bípeda pudo ser muy eficaz en estas condiciones ya que:
- Permite otear el horizonte por encima de la vegetación herbácea en busca de árboles o depredadores.
- Permite transportar cosas (como comida, palos, piedras o crías) con las manos, liberadas de la función locomotora.
- Es más lenta que la marcha cuadrúpeda, pero es menos costosa energéticamente, lo que debería ser interesante para recorrer largas distancias en la sabana, o en un hábitat más pobre en recursos que la selva.
- Expone menos superficie al sol y permite aprovechar la brisa, lo que ayuda a no recalentar el cuerpo y ahorrar agua, cosa útil en un hábitat con escasez de la misma.
Una hipótesis más elaborada ha sido propuesta por Lovejoy, y se conoce como "la tríada fundamental de la selección natural, reproducirse, comer e invertir en seguridad". Según esta hipótesis, gastar más energía en reproducirse mejor tendría un gran impacto a nivel de selección. Si una hembra pudiera tener más hijos disminuyendo los intervalos entre nacimientos y criando más de un hijo a la vez, esto revertiría en aumentar las posibilidades de dejar un mayor número de descendientes (genes) en la siguiente generación. Esto requeriría que dejara de invertir esfuerzo (energía) en buscar alimento y protegerse. Una forma de lograr esto es que los machos sean los encargados de estas tareas, tareas de acarreamiento que podría llevar a cabo al haber liberado los brazos. Sería necesario unos vínculos más estables entre machos y hembras, quizás los orígenes de una relación monógama. Esto podría explicar los caracteres sexuales secundarios tan desarrollados de nuestra especie y que repercuten en la atracción individual. Tampoco hay signos externos del estro, lo que podría ser una consecuencia de que no existe competencia entre los machos.
Hace años se argumentó también que la liberación de las manos por parte de los primeros homínidos bípedos les permitió elaborar armas de piedra para cazar, lo cual habría sido el principal motor de nuestra evolución. Hoy está claro que la liberación de las manos (que se produjo hace más de 4 millones de años) no está ligada a la fabricación de herramientas, que aconteció unos 2 millones de años después, y que los primeros homínidos no eran cazadores y que a lo sumo comían carroña esporádicamente.
4.3. Aspectos biológicos de la aparición del lenguaje simbólico.
Hablar de la aparición del lenguaje humano, lenguaje simbólico, por lógica parecería implicar que hay que hablar previamente de la cerebración, y eso es bastante cierto, pero el lenguaje humano simbólico tiene sus antecedentes en momentos y cambios morfológicos que son previos a cambios importantes en la estructura del sistema nervioso central. Por ejemplo, los chimpancés pueden realizar un esbozo primario de lenguaje simbólico basándose en la mímica (de un modo semejante a un sistema muy simple de comunicación para sordomudos).
Ahora bien, el lenguaje simbólico por excelencia es el basado en los significantes acústicos, y para que una especie tenga la capacidad de articular sonidos discretos se requieren más innovaciones morfológicas, algunas de ellas muy probablemente anteriores al desarrollo de un cerebro lo suficientemente complejo como para pensar de modo simbólico. En efecto, observemos la orofaringe y la laringe: en los mamíferos, a excepción del humano, la laringe se encuentra en la parte alta de la garganta, de modo que la epiglotis cierra la tráquea de un modo estanco al beber e ingerir comida. En cambio, en Homo sapiens, la laringe se ubica más abajo, lo que permite a las cuerdas vocales la producción de sonidos más claramente diferenciados y variados; pero al no poder ocluir completamente la epiglotis, la respiración y la ingesta deben alternarse para que el sujeto no se ahogue. El acortamiento del prognatismo, que se compensa con una elevación de la bóveda palatina, facilitan el lenguaje oral. Otro elemento de relevante importancia es la posición y estructura del hioides, su gracilidad y motilidad permitirán un lenguaje oral lo suficientemente articulado.
Estudios realizados en la Sierra de Atapuerca (España) evidencian que Homo antecessor, hace unos 800.000 años, ya tenía la capacidad, al menos en su aparato fonador, para emitir un lenguaje oral lo suficientemente articulado como para ser considerado simbólico, aunque la consuetudinaria fabricación de utensilios (por toscos que fueran) por parte del Homo habilis hace unos 2 millones de años, sugiere que en éstos ya existía un lenguaje oral articulado muy rudimentario pero lo suficientemente eficaz como para transmitir la suficiente información o enseñanza para la confección de los toscos artefactos.
Además de todas las condiciones recién mencionadas, imprescindibles para la aparición de un lenguaje simbólico, se debe hacer mención de la aparición del gen FOXP2, que resulta básico para la posibilidad de tal lenguaje y del pensar simbólico, como se verá a continuación.
 El gen FOXP2.
El gen FOXP2.
El gen y proteína FOXP2, descubierto en los años noventa del siglo XX, está relacionado con el lenguaje humano: su mutación se correlaciona con determinados trastornos específicos del lenguaje.
Aunque se ha acuñado la expresión gen del lenguaje o del habla para caracterizarlo, se trata de un factor más entre los responsables de la competencia humana para el lenguaje. No obstante, es un indicio de que, probablemente, en el lenguaje humano están involucrados factores de tipo genético: aunque se desconoce su función exacta, parece imprescindible para un desarrollo normal del mismo.
Se han barajado dos hipótesis acerca de cuál es el déficit central que provoca la anomalía en FOXP2: unos autores defienden que se trata de problemas gramaticales y otros de problemas motores (una pronunciación deficiente impediría un habla normal).
FOXP2 se expresa en varias zonas del cerebro durante la embriogénesis, aunque no está claro si la activación del gen se produce en la fase embrionaria o si eso ocurre en el momento en que se empieza a aprender a hablar. Sus niveles más altos aparecen en la capa VI del córtex, sobre todo en estructuras subcorticales de la base del cerebro (muy próximas al cuerpo calloso): núcleos basales, tálamo y cerebelo. Además, está presente en la embriogénesis de otros órganos humanos: pulmones, intestino y corazón.
Se ha sugerido, además, que la misma versión del gen estaba ya presente en los neandertales, habiendo sido modificado por selección natural durante la evolución humana reciente en los últimos 200.000 años.
No se trata de un gen exclusivo del ser humano, sino que es probable que exista en todos los vertebrados y, además, de una forma muy parecida. En lo que respecta al cerebro, se expresa en ellos en las mismas áreas que en el hombre: núcleos basales, cerebelo, tálamo y córtex o regiones equivalentes.
Se ha sugerido que en determinadas aves el gen podría contribuir a la plasticidad del canto. Por otra parte, las versiones de la proteína en el chimpancé y el ser humano difieren en dos aminoácidos, algo que ha reforzado la hipótesis de que ciertas alteraciones en el gen podrían haber impulsado la evolución del lenguaje.
El gen del lenguajePor Juan Luis Arsuaga (30 de enero de 2002) [Saltarse este artículo]

En contraste con las dificultades de los adultos portadores de ese gen para hablar correctamente, los niños normales nos sorprenden siempre por su asombrosa destreza a la hora de manejar el idioma materno.
Por eso nos hacen gracia, porque resulta chocante que un mocoso que apenas levanta un metro del suelo hable como un gramático. Los errores que cometen los críos al hablar, y que tanto nos divierten, se deben a que se pasan de listos y aplican la regla sin excepción, convirtiendo en regulares los verbos irregulares, por ejemplo. De una manera inconsciente parecen tener muy clara la estructura interna del idioma. Otras destrezas como la aritmética, aparentemente menos difíciles que aprender un idioma, las adquieren los niños más tarde, con mucho esfuerzo y un profesor. Hablar es algo natural en los niños; multiplicar no.
El lingüista Noam Chomsky llegó hace 40 años a la convicción de que, efectivamente, los niños vienen al mundo con estructuras neuronales que los capacitan para aprender un idioma; en cierto modo, disponen de un "órgano para el lenguaje". Debe de haber algo común, un conjunto de reglas, en todas las lenguas a pesar de su enorme diversidad; por eso es posible la traducción, el trasvase entre idiomas. Esa base común, sería, según Chomsky, innata.
El gen anómalo que presenta la familia investigada sólo se diferencia de su forma normal en una base de la cadena de nucleótidos (adenina en lugar de guanina) en una sola hebra de la doble hélice del ADN. Ese cambio modifica un sólo aminoácido en la cadena de la proteína para la que codifica el gen. El resultado de una variación tan minúscula en una molécula es sorprendente: un grave problema lingüístico.
¿Debe deducirse de lo descubierto que existe algo así como un "gen para el lenguaje"? Nada de eso. El que un cambio de base impida formar bien las frases no significa que ese gen concreto sea el responsable del lenguaje. Pero sí parece querer decir que hay una base genética para el lenguaje, reivindicando al lingüista Chomsky. Y si el lenguaje, que es tan importante, tan humano, y tan complejo, tiene una base biológica, ¿cuántos otros aspectos de nuestra humanidad la tienen? Hay todo un abanico de opiniones al respecto, desde los que piensan, como decía tajantemente Ortega y Gasset, que el ser humano no tiene naturaleza y todo en él es cultura, hasta los convencidos de un determinismo genético estricto de la personalidad, llegando casi hasta la ecuación "un gen, un rasgo del carácter". Entre medias se encuentran los que imaginan que los genes establecen predisposiciones muy generales, amplias avenidas que recorremos en nuestra infancia y a lo largo de las cuales se va formando la mente del adulto en diálogo constante con el ambiente cultural en el que se produce el desarrollo. El descubrimiento del gen citado en el cromosoma 7 no soluciona nuestras dudas, pero puede ser un primer paso, una primera aportación del Proyecto Genoma Humano al conocimiento de las bases biológicas del comportamiento. Y también representa quizás el nacimiento de una nueva disciplina científica: la genética cognitiva.
Audición y lenguajePor José María Bermúdez de Castro (3 de diciembre de 2008) [Saltarse este artículo]

Durante algunos años, mi compañero de proyecto Ignacio Martínez dedicó mucho esfuerzo a investigar el lenguaje de los homininos de la Sima de los Huesos de Atapuerca. Estos homininos tienen medio millón de años de antigüedad y no están relacionados de manera directa con nosotros, sino con los neandertales. Ante la dificultad de llegar a conclusiones satisfactorias que convencieran a los especialistas que examinaban sus investigaciones, Martínez tuvo una idea brillante: ¿por qué no estudiar la audición de estos homininos? Para ello contaba con fósiles jamás hallados en ningún otro yacimiento: varios ejemplares de los huesecillos del oído, martillo, yunque y estribo, así como con muchos datos sobre otras variables de oído de estos homininos. Los restos del yacimiento están embebidos en una arcilla finísima, que ha permitido la perfecta conservación de regiones anatómicas poco frecuentes en el registro fósil.
Ayudado por especialistas en ingeniería acústica, Ignacio Martínez pudo construir un modelo para determinar la curva de audición de los homininos de Atapuerca. Si los chimpancés tienen dos picos de máxima audición a 1 y 8 kilo-hertzios (KHz), como adaptación específica a su medio, los humanos actuales tenemos un curva con máxima audición entre 2 y 4 KHz. Los resultados del estudio revelaron que los homininos de la Sima de los Huesos tenían también su máxima audición entre 2 y 4 KHz.
Lo más interesante de este estudio es que nuestro lenguaje se emite precisamente en la banda de entre 2 y 4 KHz. En otras palabras, desde hace al menos 600.000 años y tal vez desde hace mucho más tiempo, una especie distinta de la nuestra (Homo heidelbergensis) se comunicaba a corta distancia mediante sonidos que tenían la misma frecuencia que nuestro lenguaje actual. Esto no significa haber demostrado que aquellos homininos de Atapuerca hablaran igual que nosotros, pero sí que utilizaban una comunicación parecida a la nuestra para transmitir información, ideas y conceptos, que se pueden inferir del estudio del registro arqueológico.4.4. Cerebración.
La cerebración, tanto como la corticalización, son fenómenos biológicos muy anteriores a la aparición de los homínidos. Sin embargo en éstos, y en especial en Homo sapiens, la cerebración y la corticalización adquieren un grado superlativo.
El cerebro de Homo sapiens, en relación a la masa corporal, es uno de los mayores. Más llamativo es el consumo de energía metabólica que requiere el cerebro: un 20% de toda la energía corporal, y aún cuando la longitud de los intestinos humanos evidencian los problemas que se le presentan.
En Homo sapiens el volumen oscila entre los 1.200 a 1.400 cm3, siendo el promedio global actual de 1.350 cm3. Sin embargo no basta un incremento del volumen, sino cómo se dispone, esto es, cómo está dispuesta la estructura del sistema nervioso central y del cerebro en particular. Por término medio, los Homo neanderthalensis pudieron haber tenido un cerebro de mayor tamaño que el de nuestra especie, pero la morfología de su cráneo demuestra que la estructura cerebral era muy diferente: con escasa frente, los neandertales tenían poco desarrollados los lóbulos prefrontales y, en especial, muy poco desarrollados los lóbulos frontales. El cráneo de Homo sapiens no sólo tiene una frente adelantada (sobre el rostro), sino que es también más alto en el occipucio (cráneo muy abovedado), lo que permite el desarrollo de los lóbulos prefrontales y frontales. De todos los mamíferos, Homo sapiens es el único que tiene la faz ubicada bajo los lóbulos frontales.

Se ha hecho mención en el apartado dedicado a la aparición del lenguaje articulado de la importancia del gen FOXP2. Dicho gen es el encargado del desarrollo de las áreas del lenguaje y de las áreas de síntesis (las áreas de síntesis se encuentran en el córtex de los lóbulos prefrontales y, sobre todo, frontales). El aumento del cerebro y su especialización permitió la aparición de la llamada lateralización, o sea, una diferencia muy importante entre el hemisferio izquierdo y el hemisferio derecho del cerebro. El hemisferio izquierdo tiene desarrollado en su córtex áreas específicas que posibilitan el lenguaje simbólico basado en significantes acústicos: el área de Wernicke y el área de Broca.
Es casi seguro que ya hace 200.000 años los sujetos de la especie Homo sapiens tenían un potencial intelectual equivalente al de la actualidad, pero para que se activara tal potencial tardaron milenios: el primer registro de conducta artística conocido se data hace sólo unos 75.000 años, los primeros grafismos y expresiones netamente simbólicas fuera del lenguaje hablado se datan hace sólo entre 40.000 y 35.000 años, las primeras escrituras datan de hace entre 5.500 ó 5.000 años, en el valle del Nilo ó en la Mesopotamia asiática.
Se ha dicho, también líneas antes, que Homo sapiens mantiene características de estructura craneal neoténicas, ya que recuerdan a las de un chimpancé infantil. En efecto, tal morfología es la que permite tener la frente sobre el rostro y los lóbulos frontales desarrollados. La cabeza de Homo sapiens, para contener tal cerebro, es muy grande, aún en el feto y en el neonato, razón principal por la cual los partos son difíciles, sumada a la disposición de la pelvis.
Una solución parcial a esto es la heterocronía: el neonato humano está muy incompletamente desarrollado en el momento del parto. Puede decirse (con algo de metáfora) que la gestación en el ser humano no se restringe a los ya de por sí prolongados nueve meses intrauterinos, sino que se prolonga extrauterinamente hasta, al menos, los cuatro primeros años. En efecto, el infante está completamente desvalido durante años, tanto es así que entre los 2 a 4 años es cuando tiene lo suficientemente desarrolladas las áreas visuales del cerebro como para tener una percepción visual de su propio ser (estadio del espejo descubierto por Jacques Lacan en la década de 1930). Ahora bien, si Homo sapiens tarda mucho en poder tener una percepción plena de su imagen corporal es interesante saber que es uno de los pocos animales que se percibe al ver su imagen reflejada (sólo se nota esta capacidad en bonobos, chimpancés, y si acaso en gorilas, orangutanes, delfines y elefantes).
Tal es la prematuración de Homo sapiens, que mientras un chimpancé neonato tiene una capacidad cerebral de un 65% de la de un chimpancé adulto, o la capacidad de Australopithecus afarensis era en el parto de un 50% respecto a la de su edad adulta, en un Homo sapiens neonato tal capacidad no supera al 25% de la capacidad que tendrá a los 45 años (que es aproximadamente cuando se desarrolla totalmente el cerebro humano).
|

| Ver vídeo complementario sobre la cerebración durante el proceso de hominización [3:35] |
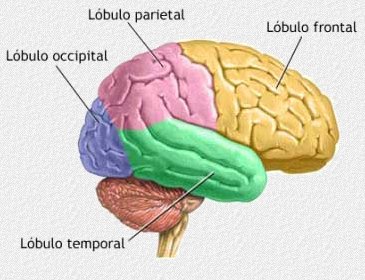
4.4.1. Estructura del cerebro.
El cerebro humano tiene una estructura distinta a la de otros primates y en la superficie cerebral podemos localizar regiones que participan de forma especializada en diferentes actividades físicas o cognitivas. Debido a la especialización de las diferentes regiones del cerebro podemos observar grandes diferencias morfológicas en la estructura del cerebro de los humanos y los chimpancés.
En los fósiles, la estructura del cerebro puede ser estudiada a través de las impresiones que dejan en las paredes internas del cráneo los sulcos, circunvoluciones, cisuras y venas meníngeas. En muchos casos estas impresiones son muy tenues y limitan nuestro estudio únicamente a la superficie del cerebro.
En los Australopithecus y Paranthropus existe disparidad de opiniones sobre el grado de reorganización cerebral que tienen respecto de los antropomorfos. Sin embargo, todos los autores están de acuerdo en que Homo habilis y Homo ergaster ya tienen una morfología cerebral similar a la de humanos modernos y presentan las siguientes características:
- Asimetría entre los hemisferios.
- Desarrollo y complejidad del lóbulo frontal.
El cerebro humano actual es muy asimétrico. La asimetría entre ambos hemisferios cerebrales esta relacionada con la lateralización cerebral, es decir, la especialización de cada una de las regiones cerebrales en distintas funciones. En los cráneos de los primeros Homo ya puede observarse esta asimetría cerebral.
4.4.2. El desarrollo del lóbulo frontal.
El lóbulo frontal es responsable de algunas capacidades cognitivas exclusivas de los humanos o que en éstos están mucho más desarrolladas. Entre las funciones del lóbulo frontal se encuentran la de establecer la secuencia de movimientos del aparato fonador, el control de las emociones, la concentración, la planificación y anticipación, el control de la memoria. Como ya se ha dicho, los neandertales tenían más volumen encefálico, pero su lóbulo frontal estaba mucho menos desarrollado.
A lo largo de la evolución humana, el lóbulo frontal ha crecido en tamaño absoluto y en relación al resto del cerebro. Además, su superficie se ha hecho más compleja aumentando el número de sulcos en su superficie.