Los fósiles más antiguos de Homo sapiens (los anteriormente mencionados Omo I y II) tienen, como se ha dicho, una antigüedad de casi 200.000 años y proceden del sur de Etiopía (formación Kibish del río Omo), considerada como la cuna de la humanidad. A estos restos fósiles siguen en antigüedad los de Homo sapiens idaltu, con 160.000 años.
Algunos datos de genética molecular, concordantes con hallazgos paleontológicos, sostienen que todos los seres humanos descienden de una misma "Eva mitocondrial"; esto quiere decir que, según los rastreos del ADN mitocondrial (que sólo se transmite por vía materna), toda la humanidad actual tiene una antecesora común que habría vivido en el noreste de África (en territorios que corresponden a los actuales estados de Etiopía y Kenia) hace entre 140.000 y 200.000 años. En otros estudios sin embargo, basados en el polimorfismo del complejo mayor de histocompatibilidad, se sugiere que en los últimos millones de años la especie humana o sus ancestros jamás pudieron haber comprendido menos de 100.000 individuos, lo que derrumba la teoría del cuello de botella del ADN mitocondrial y la Eva ancestral. Estas conclusiones son derivadas del hecho de que humanos y chimpancés comparten muchas diferentes variedades de los mismos genes que no pudieron haberse trasmitido de una especie a otra si hubiera existido sólo un humano originario.
Otros indicios derivados de recientes investigaciones sugieren que la de por sí exigua población de Homo sapiens hace unos 74.000 años se redujo al borde de la extinción al producirse el estallido del volcán Toba, según la teoría de la catástrofe de Toba. Si esto es cierto, significaría que el pool genético de la especie se habría restringido de tal modo que se habría potenciado la unidad genética de la especie humana.
 Teoría de la catástrofe de Toba
Teoría de la catástrofe de Toba
La teoría de la catástrofe de Toba, propuesta en 1998 por Stanley H. Ambrose, establece que la evolución humana fue afectada por un reciente evento de tipo volcánico. Este evento habría reducido la población mundial a 10.000 o incluso unas 1.000 parejas reproductoras.
La teoría catastrófica de Toba establece que una inmensa erupción volcánica cambió el curso de la historia al producir la casi extinción de la población humana (este tipo de eventos se denomina «cuello de botella poblacional»). Hace entre 70.000 y 75.000 años, el supervolcán del lago Toba, en el norte de la isla indonesia de Sumatra, explosionó como una caldera con una fuerza 3.000 veces superior a la erupción del monte Santa Helena, dejando como rastro el actual lago Toba (el lago volcánico más grande del mundo, de 100 kilómetros de largo, 30 de ancho y 505 metros de profundidad). Se han encontrado restos de esta explosión en lugares tan remotos como India.
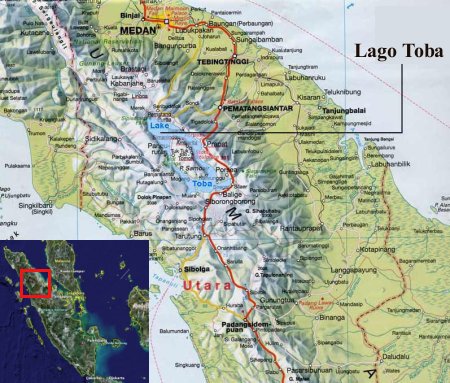
Según Ambrose, esto provocó una caída de la media de las temperaturas de 3-3,5oC, con un invierno volcánico global que pudo durar entre 6 y 7 años. En las regiones templadas produjo una disminución de las temperaturas globales de 15oC de promedio, lo que representa un cambio drástico en el ambiente, que debió producir múltiples cuellos de botella de población en varias especies humanas que debían existir en la época (Homo sapiens en África, Oriente Medio y Asia, Homo erectus y Homo floresiensis en Indonesia y Homo neanderthalensis en Europa). Este cambio aceleró a su vez la diferenciación de las poblaciones humanas aisladas, conduciendo finalmente a la extinción de todas las especies humanas (menos una, de la cual descienden los humanos actuales).
Una combinación de evidencias geológicas y modelos computacionales apoya la factibilidad de la teoría de la catástrofe de Toba, y la evidencia genética sugiere que todos los humanos actuales, a pesar de la aparente variedad, provienen de un mismo tronco formado por una población muy pequeña. Utilizando las tasas promedio de mutación genética algunos genetistas han estimado que esta población humana original vivió en una época que concuerda con el evento de Toba.3.1. Origen del hombre moderno.
Los paleoantropólogos siguen debatiendo sobre dónde evolucionó el hombre moderno y cómo se dispersó por el mundo. La cuestión es si la evolución tuvo lugar en una pequeña región de África o en una amplia área de África y Eurasia. Y aún más, si las poblaciones humanas modernas procedentes de África desplazaron a todas las poblaciones humanas allí existentes, llegando a producir en último término su extinción.
Aquellos que piensan que el hombre moderno tuvo su origen sólo en África para más tarde dispersarse por el mundo apoyan la denominada hipótesis de la emigración de África. Aquellos que piensan que el hombre moderno evolucionó por toda una gran región de Eurasia y África apoyan la denominada hipótesis multirregional.
Los investigadores han realizado numerosos estudios genéticos y han valorado cuidadosamente los fósiles para determinar cuál de estas hipótesis coincide más con la evidencia científica. Los resultados de esta investigación no confirman ni rechazan totalmente ninguna de ellas. Por lo tanto, algunos científicos piensan que la mejor explicación es un compromiso entre ambas hipótesis. El debate entre estas teorías tiene implicaciones en cuanto a qué entienden los científicos como raza humana. La cuestión que se plantea es si en el hombre moderno las diferencias físicas evolucionaron profundamente en el pasado lejano o lo hicieron en un pasado relativamente reciente.
3.1.1. La hipótesis de la emigración de África.
De acuerdo con la hipótesis de la emigración de África, también conocida como la hipótesis de sustitución o Out of Africa, las primeras poblaciones de hombres modernos procedentes de África migraron a otras regiones y sustituyeron por completo a las poblaciones hasta entonces existentes de homininos primitivos. Entre las poblaciones sustituidas se incluye al Hombre de Neandertal y a cualquier grupo superviviente de Homo erectus. Los defensores de esta hipótesis afirman que muchas características del esqueleto del hombre moderno evolucionaron en un pasado relativamente reciente (hace unos 200.000 años aproximadamente) lo que sugiere un origen único y común. Además, las similitudes anatómicas comunes a todas las poblaciones de hombres modernos superan de lejos las compartidas entre el hombre moderno y el hombre premoderno dentro de regiones geográficas determinadas. Numerosos estudios biológicos indican que la mayoría de las especies nuevas de organismos, incluidos los mamíferos, han surgido de poblaciones pequeñas y geográficamente aisladas.
3.1.2. La hipótesis multirregional.
Según la hipótesis multirregional, también conocida como la hipótesis de continuidad, la evolución del hombre moderno comenzó cuando el Homo erectus se dispersó por gran parte de Eurasia hace un millón de años. Las poblaciones regionales retuvieron algunas características anatómicas únicas durante cientos de miles de años, pero ellos también se mezclaron con poblaciones de las regiones colindantes, intercambiando rasgos hereditarios entre sí. Este intercambio de rasgos se conoce como flujo genético.
A través del flujo genético las poblaciones de Homo erectus transmitieron una serie de características cada vez más modernas, tales como un mayor tamaño de cerebro, por todo su radio geográfico. Gradualmente esto habría dado lugar a la evolución de un hominino con aspecto más moderno en toda África y Eurasia. En este caso las diferencias físicas entre los hombres actualmente serían el resultado de cientos de miles de años de evolución regional. Este es el concepto de continuidad. Así, por ejemplo, las poblaciones modernas del este asiático tienen algunas características craneanas observadas también por los científicos en fósiles del Homo erectus de esta región.
3.1.3. Contrastando ambas hipótesis.
Los científicos han contrastado las hipótesis de la emigración de África y la multirregional tanto con las evidencias genéticas como con los fósiles encontrados. Aunque los resultados no apoyan definitivamente ninguna de las dos hipótesis, se inclinan a favor de la primera.
3.1.3.1. Evidencias genéticas.
Los genetistas han estudiado las diferencias en el ADN de diferentes poblaciones humanas y calculado la velocidad previsible para que se produzca una mutación a lo largo del tiempo. Dividiendo el número total de diferencias genéticas entre dos poblaciones por la velocidad esperada de mutación se obtiene una estimación del momento en el que ambas poblaciones compartieron un antepasado común. Muchas estimaciones de antepasados evolutivos se basan en estudios del ADN mitocondrial. A diferencia del ADN nuclear, la herencia mitocondrial pasa sólo de la madre a su descendencia. El ADN mitocondrial también acumula mutaciones con una rapidez diez veces mayor que el ADN nuclear. Su estructura cambia tan rápidamente que los científicos pueden medir fácilmente las diferencias entre una población humana y otra. Dos poblaciones estrechamente relacionadas deberían tener sólo pequeñas diferencias de ADN mitocondrial. Por el contrario, dos poblaciones muy lejanamente relacionadas entre sí deberían presentar grandes diferencias.
Las investigaciones genéticas para el conocimiento de los orígenes del hombre moderno han dado lugar a dos importantes descubrimientos. En primer lugar, las variaciones del ADN mitocondrial entre diferentes poblaciones humanas son mínimas en comparación con las de otras especies de animales. Esto significa que todo el ADN mitocondrial humano se originó a partir de un linaje ancestral único en un pasado bastante reciente. La mayoría de las estimaciones de la velocidad de mutación del ADN mitocondrial sugieren que este antepasado femenino vivió hace unos 200.000 años. Además, el ADN mitocondrial de las poblaciones del continente africano varía más que el de los pueblos de otros continentes. Esto sugiere que su ADN mitocondrial ha ido cambiando durante un periodo de tiempo más largo que el de las poblaciones de cualquier otra región, y que todos los seres humanos actuales heredaron su ADN mitocondrial de una mujer africana, que a veces recibe el nombre de Eva mitocondrial. A partir de esta evidencia algunos genetistas y antropólogos han llegado a la conclusión de que el hombre moderno tuvo su origen en una población pequeña de África para posteriormente dispersarse desde allí.
Sin embargo, los estudios del ADN mitocondrial presentan, entre otros, cuatro puntos débiles principales:
- La velocidad estimada de mutación del ADN mitocondrial varía de un estudio a otro y algunas estimaciones señalan la fecha de origen hace unos 850.000 años, la era del Homo erectus.
- El ADN mitocondrial es sólo una pequeña parte del material genético total que hereda el ser humano, ya que el resto de nuestro material genético (aproximadamente unas 400.000 veces mayor que la cantidad de ADN mitocondrial) procede de muchos individuos que vivían en la misma época de la Eva africana, posiblemente en muchas regiones diferentes.
- El momento en el que el ADN mitocondrial moderno comenzó a diversificarse no coincide necesariamente con el origen de los rasgos biológicos y habilidades culturales del hombre moderno.
- La menor diversidad del ADN mitocondrial moderno fuera de África puede proceder de épocas en las que se produjo una reducción de las poblaciones europeas y asiáticas tal vez debido a cambios climáticos.
A pesar de estas críticas, muchos genetistas continúan apoyando la hipótesis de emigración de África para el origen del hombre moderno. Además, algunos estudios del ADN nuclear también sugieren un origen africano para una serie de genes.
3.1.3.2. Evidencias fósiles.
Al igual que ocurre con las evidencias genéticas, las evidencias fósiles tampoco apoyan ni rechazan totalmente ninguna de las hipótesis sobre los orígenes del hombre moderno, aunque muchos científicos piensan que favorecen el origen africano del Homo sapiens durante los últimos 200.000 años. Los cráneos más antiguos con aspecto moderno conocidos proceden de África y datan de hace aproximadamente unos 200.000 años. El siguiente fósil en antigüedad fuera de África procede de Oriente Próximo y está datado en unos 90.000 años de antigüedad. No existen fósiles de hombres modernos en Europa de más de 40.000 años. Además, el hominino moderno europeo (a menudo denominado Hombre de Cro-Magnon) tenía los huesos de los miembros inferiores alargados, como las poblaciones africanas que estaban adaptadas a climas cálidos tropicales. Esto sugiere que las poblaciones de regiones más cálidas sustituyeron a las existentes en regiones europeas más frías, tales como los neandertales.
Los fósiles también muestran que las poblaciones de homininos modernos vivían al mismo tiempo y en las mismas regiones que las poblaciones de neandertales y Homo erectus, pero que cada una de ellas mantuvo unas características físicas diferenciales. Los diferentes grupos se solaparon en Oriente Próximo y en el Sudeste Asiático durante unos 30.000 a 50.000 años. El mantenimiento de diferencias físicas durante todo este tiempo implica que los homininos primitivos y modernos, por lo general, no podían o no llegaron a mezclarse.
Por otro lado, los fósiles de hombres modernos y homininos primitivos en algunas regiones muestran una continuidad de determinadas características físicas. Estas similitudes podrían indicar una evolución multirregional. Así, por ejemplo, tanto los cráneos primitivos como los cráneos modernos del este de Asia tienen pómulos y huesos nasales menos pronunciados que los de otras regiones. Por el contrario, las mismas partes del rostro sobresalen más en los cráneos de los humanos primitivos y del hombre moderno europeo. Asumiendo que estos rasgos estuvieron determinados más por la herencia genética que por los factores ambientales, un hominino primitivo pudo haber dado origen al hombre moderno en algunas regiones o al menos haberse cruzado con emigrantes de aspecto moderno.
3.1.4. Una hipótesis de compromiso.
Cada una de las hipótesis en litigio sobre los orígenes del hombre moderno tiene sus puntos fuertes y sus puntos débiles. La evidencia genética parece apoyar la hipótesis de la emigración de África. En la mitad occidental de Eurasia y en África esta hipótesis también parece ser la mejor explicación, especialmente con respecto a la aparente sustitución del neandertal por poblaciones modernas. Por otro lado, la hipótesis multirregional parece explicar en parte la continuidad regional encontrada en las poblaciones de Asia Oriental.
Por esta razón, muchos paleoantropólogos defienden una teoría sobre los orígenes del hombre moderno que combina elementos de la hipótesis de la emigración de África y elementos de la hipótesis multirregional. Un hominino con rasgos modernos podría haber surgido por primera vez en África o haberse unido allí, como resultado del flujo genético, con poblaciones de otras regiones. Estas poblaciones africanas pueden a continuación haber sustituido al hominino primitivo en determinadas regiones, tales como el oeste de Europa y Oriente Próximo. Pero en los demás lugares, especialmente en Asia Oriental, puede haber tenido lugar un flujo genético entre las poblaciones locales de homininos primitivos y modernos que hubiese dado lugar a características regionales diferenciadoras y permanentes.
Estas tres hipótesis (las dos en litigio y la de compromiso) reconocen la fuerte unidad biológica de todos los hombres. En la hipótesis multirregional, esta unidad es el resultado de cientos de miles de años de flujo genético continuo entre todas las poblaciones humanas. Por otra parte, y de acuerdo con la hipótesis de la emigración de África, las similitudes entre las poblaciones humanas actuales resultan de un origen común reciente. La postura de compromiso acepta ambas hipótesis como explicaciones razonables y compatibles sobre los orígenes del hombre moderno.
 Orígenes de los humanos modernos: ¿Multirregional o Out of Africa?
Orígenes de los humanos modernos: ¿Multirregional o Out of Africa?Por Donald Johanson (mayo de 2001) [Saltarse este artículo]

Entendiendo el problema
El Modelo de la Continuidad Multirregional15 contiende que después de que Homo erectus salió de África y se dispersó a otras partes del Viejo Mundo, las poblaciones regionales evolucionaron lentamente en los humanos modernos. Este modelo tiene los siguientes componentes:
- Algún nivel de flujo genético entre las poblaciones separadas geográficamente previno la especiación después de la dispersión.
- Todos los humanos que viven hoy en día derivaron de la especie Homo erectus que salió de África hace casi dos millones de años.
- La selección natural, actuando sobre las poblaciones regionales desde su dispersión original, es responsable de las variantes regionales (a veces llamadas razas) que vemos hoy en día.
- La aparición de Homo sapiens no se restringió a un área solamente, sino que fue un fenómeno que ocurrió a través del rango geográfico completo donde vivían los humanos.
En contraste, el Modelo Out of Africa13 afirma que los humanos modernos evolucionaron de forma relativamente reciente en África, migraron a Eurasia y reemplazaron a todas las poblaciones que habían descendido de Homo erectus. Los siguientes principios son críticos en este modelo:
- Después de que Homo erectus migró fuera de África, las diferentes poblaciones se aislaron reproductivamente, evolucionando independientemente y, en algunos casos como en los neandertales, en especies separadas; Homo sapiens se originó en un sólo sitio, probablemente en África (geográficamente esto incluye Oriente Medio).
- Homo sapiens migró finalmente fuera de África y reemplazó a todas las otras poblaciones humanas, sin entrecruzarse con ellas.
- La variación en los humanos modernos es un fenómeno relativamente reciente.
La perspectiva multirregional postula que los genes de todas las poblaciones humanas del Viejo Mundo fluyeron entre regiones diferentes y se mezclaron, contribuyendo a lo que observamos actualmente en los humanos modernos. La hipótesis del reemplazo sugiere que los todos genes de los humanos completamente modernos vinieron de África. A medida que estas gentes migraron, reemplazaron a todas las otras poblaciones humanas, con muy poco o ningún entrecruzamiento.
Para entender la controversia, necesitamos evaluar la evidencia anatómica, arqueológica y genética.
La evidencia anatómica
En una época antes de hace un millón de años, los primeros homínidos, a veces llamados Homo ergaster, salieron de África y se dispersaron a otras partes del Viejo Mundo. Al vivir en áreas geográficas dispares, su morfología se diversificó a través de los procesos de la deriva genética y de la selección natural.
- En Asia estos homínidos evolucionaron en el Hombre de Pekín y en el Hombre de Java, referidos en conjunto como Homo erectus.
- En Europa y en Asia occidental evolucionaron en los neandertales.
Los neandertales vivieron en un aislamiento casi total en Europa durante un período relativamente frío, el cual hasta incluyó glaciaciones. Los neandertales se distinguen por un juego de características anatómicas únicas, incluyendo:
- Una cavidad craneal grande, larga y baja, con una protuberancia frontal bien desarrollada y doblemente arqueada.
- Un esqueleto facial masivo con una cara media bien proyectada, mejillas echadas hacia atrás y apertura nasal grande, con senos nasales también grandes.
- Una extraña región occipital del cráneo con un saliente o "chichón".
- Molares con cámaras pulpares agrandadas y con dientes incisivos grandes y a menudo altamente desgastados.
- Una mandíbula sin barbilla y con una gran apertura detrás del último molar.
- Un tórax masivo, con los brazos anteriores y las pantorrillas relativamente cortas.
- A pesar de ser bajos de estatura, poseían esqueletos muy robustos, con huesos de pared muy gruesa.
- Clavículas largas y escápulas muy anchas.
Para hace 130.000 años, y seguido de un periodo prolongado de evolución independiente en Europa, los neandertales eran anatómicamente tan únicos que es mejor clasificarlos como una especie diferente: Homo neanderthalensis. Este es un ejemplo clásico de aislamiento geográfico que llevó finalmente a un evento de especiación.
En contraste, aproximadamente al mismo tiempo en África, un plan corporal esencialmente como el nuestro había aparecido. A pesar de que estos primeros Homo sapiens eran anatómicamente modernos, no lo eran en su comportamiento. Es muy importante anotar que la anatomía moderna evolucionó antes que el comportamiento moderno; estos primeros sapiens estaban caracterizados por lo siguiente:
- Una cavidad craneal con una frente vertical, un occipital redondeado y una protuberancia frontal reducida.
- Un esqueleto facial reducido sin una cara media proyectada hacia adelante.
- Una mandíbula con barbilla.
- Un esqueleto moderno y menos robusto.
De ahí, la evidencia anatómica y paleogeográfica sugiere que los neandertales y los humanos modernos estuvieron aislados los unos de los otros y que evolucionaron separadamente en dos especies diferentes.
La evidencia arqueológica
De manera muy interesante, mientras que los neandertales y los primeros Homo sapiens se distinguían los unos de los otros por un juego de características anatómicas obvias, arqueológicamente eran muy similares. Los homínidos africanos de la Edad de Piedra Media (Homo sapiens) y sus contemporáneos neandertales europeos del Paleolítico Medio poseían juegos de artefactos caracterizados de la siguiente manera:
- Poca variación en tipos de herramientas líticas, con una preponderancia de herramientas de lascas de piedra difíciles de clasificar en categorías discretas.
- Los juegos de herramientas eran muy similares a lo largo de grandes extensiones de tiempo y de distancia.
- Una virtual falta de herramientas hechas de hueso, asta o marfil.
- Los entierros no tenían bienes funerarios o evidencia de ceremonias o rituales.
- La cacería estaba generalmente limitada a especies menos peligrosas y no hay evidencia de pesca.
- Las densidades poblacionales eran aparentemente bajas.
- No hay evidencia de estructuras de vivienda y las fogatas eran rudimentarias.
- Tampoco hay evidencia de arte o decoración.
El cuadro arqueológico cambió dramáticamente cerca de los 40.000 a 50.000 años atrás, con la aparición de los humanos de comportamiento moderno. Esto representó un cambio abrupto y dramático en los patrones de subsistencia, en las herramientas y en la expresión simbólica. El tremendo cambio en la adaptación cultural no fue solamente un cambio cuantitativo, sino que representó una separación significativa del comportamiento humano anterior, reflejando una gran transformación cualitativa. Fue literalmente una "explosión creativa" la cual exhibió la "ingenuidad tecnológica, las formaciones sociales y la complejidad ideológica de los cazadores-recolectores históricos".7 Esta revolución humana fue precisamente la que nos hizo lo que somos hoy.
La aparición del comportamiento humano moderno completo aparentemente ocurrió en África antes de ocurrir en otras partes del Viejo Mundo, pero se dispersó rápidamente debido a los movimientos de las poblaciones a otras regiones geográficas. El estilo de vida del Paleolítico Superior, como fue llamado, era esencialmente el de cazar y recolectar. Esta adaptación cultural fue tan exitosa que, hasta hace aproximadamente unos 11.000 años, los homínidos en todo el mundo subsistían esencialmente como cazadores-recolectores.
En el Paleolítico Superior de Eurasia, o en la Edad de Piedra Tardía como se le conoce en África, la evidencia arqueológica se distingue fuertemente al ser contrastada con aquella del Paleolítico Medio o Edad de Piedra Media. Estuvo caracterizada por innovaciones significativas:
- Una diversidad asombrosa en los tipos de herramientas líticas.
- Los tipos de herramientas cambian significativamente en el espacio y en el tiempo.
- Los artefactos eran manufacturados en hueso, asta y marfil además de la piedra.
- Los artefactos de piedra eran manufacturados principalmente en hojas y se pueden clasificar fácilmente en categorías discretas, presumiblemente reflejando usos especializados.
- Los entierros iban acompañados de ceremonias rituales y contenían una rica diversidad de bienes funerarios.
- Las viviendas y fogatas eran bien diseñadas y construidas.
- La cacería de animales peligrosos y la pesca se daban con regularidad.
- La densidad de las poblaciones era mayor.
- El arte era abundante y elaborado, así como también el uso generalizado de adornos personales.
- Las materias primas, como lascas de piedra y conchas, eran objeto de comercio a través de grandes distancias.
Los Homo sapiens del Paleolítico Superior o Edad de Piedra Tardía fueron primordialmente modernos en apariencia y en comportamiento. Aún no se explica con precisión cómo ocurrió esta transformación, pero aparentemente se restringió a los Homo sapiens y no ocurrió en los neandertales. Algunos arqueólogos han invocado una explicación comportamental para explicar el cambio. Por ejemplo, Soffer11 sugiere que cambios en las relaciones sociales, tales como el desarrollo de la familia nuclear, jugaron un papel clave en la transformación.
Por su parte, Klein7 ofrece la noción de que probablemente un cambio biológico causado por mutaciones jugó el papel clave en la emergencia de los humanos de comportamiento moderno. Su explicación basada en la biología implica que ocurrió una reorganización neural mayor en el cerebro, trayendo consigo una mejora significativa en la manera en que el cerebro procesaba la información. Esta hipótesis es muy difícil de corroborar ya que los cerebros no se fosilizan. Sin embargo, es significativo que no se observan cambios en la forma de los cráneos entre los Homo sapiens tempranos y los tardíos. El registro arqueológico, con su abundante evidencia de artes y rituales, sólo nos permite concluir que estas gentes del Paleolítico Superior o Edad de Piedra Tardía poseían habilidades lingüísticas equivalentes a las nuestras. Para muchos antropólogos esto representa un salto evolutivo final hacia la modernidad completa.
Poco después de la llegada a Europa de los humanos completamente modernos, los neandertales comenzaron a declinar rápidamente, hasta culminar en su desaparición hace aproximadamente 30.000 años. Aparentemente, los neandertales no pudieron sobrevivir a los tecnológicamente avanzados humanos modernos que invadieron Europa. La evidencia del entrecruzamiento entre estos dos tipos es dudosa.
Evidencia genética
La investigación sobre los patrones de variación genética en las poblaciones humanas modernas apoya la perspectiva de que el origen del Homo sapiens es el resultado de un evento reciente, consistente con el Modelo Out of Africa.
- Los estudios en el ADN contemporáneo, especialmente del ADN mitocondrial (ADNm) el cual sólo está presente en los orgánulos celulares llamados mitocondrias, revelan que los humanos son sorprendentemente homogéneos, con una variación genética relativamente pequeña.1,5
- El alto grado de similitud entre las poblaciones humanas contrasta fuertemente con la condición observada en los chimpancés, nuestros parientes vivos más cercanos.2 De hecho, existe una variación genética significativamente mayor entre dos individuos de chimpancés tomados de la misma población que la que hay entre dos humanos tomados al azar de una misma población. Más aún, la variación genética entre las poblaciones de chimpancés es inmensamente mayor que las diferencias entre las poblaciones humanas de Europa, Asia y África.
- En apoyo al origen africano para Homo sapiens, el trabajo de Cann y Wilson1 ha demostrado que el nivel más alto de variación genética en el ADNm ocurre en poblaciones africanas. Esto implica que el Homo sapiens apareció primero en África y desde ese entonces tuvo un periodo de tiempo largo para acumular diversidad genética. Más aún, usando la distancia genética entre las poblaciones de África y otras poblaciones como una medida del tiempo, sugieren que Homo sapiens surgió entre hace 100.000 y 400.000 años.
- La pequeña cantidad de variación genética en las poblaciones humanas modernas sugiere que nuestros orígenes pueden reflejar una población fundadora de Homo sapiens relativamente pequeña. El análisis del ADNm hecho por Rogers y Harpending12 apoya la perspectiva de que una población pequeña de Homo sapiens, de entre 10.000 a 50.000 individuos, salió de África durante el período aproximado de 50.000 a 100.000 años atrás.
Los científicos han tenido éxito reciente en extraer el ADN de varios esqueletos de neandertales.8 Después de un cuidadoso análisis, particularmente del ADNm pero también ahora de una cantidad de ADN nuclear, es aparente que el ADN de los neandertales es muy diferente del nuestro. Al medir el grado de diferencia entre el ADN de los neandertales y el de los humanos modernos, los autores sugieren que estos dos linajes se separaron hace más de 400.000 años.
A pesar de estar aún en su infancia, este tipo de estudio genético apoya la perspectiva de que los neandertales no se entrecruzaron con los Homo sapiens que migraron a Europa. Es entonces muy probable que los humanos modernos no lleven en su ADN genes de los neandertales.
Consideraciones adicionales
La cronología en Oriente Medio no apoya al Modelo de Multirregionalidad, donde los neandertales y los humanos anatómicamente modernos se solaparon durante un largo periodo de tiempo.
- Algunas cavernas en Israel, más notablemente las de Qafzed y Skhul, están fechadas en casi 100.000 años y contienen esqueletos de humanos anatómicamente modernos. Más aún, se conocen restos de neandertales provenientes de sitios como la caverna de Tabun, de 110.000 años de edad, la cual precede por unos 10.000 años en la región al Homo sapiens más temprano.
- La presencia de neandertales en otras dos cuevas en Israel, Amud y Kebara, fechadas aproximadamente en 55.000 años, significa que los neandertales y los Homo sapiens se solaparon en la región durante por lo menos 55.000 años. Si los Homo sapiens estuvieron en esta región por unos 55.000 años antes de la desaparición de los neandertales, entonces no hay ninguna razón para asumir que los neandertales evolucionaron en los humanos modernos.
- La evidencia arqueológica de Europa sugiere que los neandertales pueden haber sobrevivido en la Península Ibérica hasta quizás tan recientemente como hace 30.000 a 35.000 años. Los humanos completamente modernos aparecieron por primera vez en Europa hace entre 35.000 y los 40.000 años, trayendo con ellos la tradición de herramientas del Paleolítico Superior conocida como Auriñaciense. De allí, los neandertales y los humanos completamente modernos pueden haberse solapado en Europa durante un tiempo de hasta 10.000 años. De nuevo, con los humanos completamente modernos en la escena, no es necesario tener a los neandertales evolucionando hacia los humanos modernos, lo cual le da más peso a la perspectiva de que los humanos reemplazaron a los neandertales.
- Sin embargo, la situación en el Sur de Francia no es tan clara. Aquí, en varios sitios fechados en 40.000 años aproximadamente, existe evidencia de una industria arqueológica conocida como Châtelperroniense, la cual contiene elementos de artefactos del Paleolítico Medio y Alto. Los homínidos de estos sitios son claramente neandertales, encendiendo las especulaciones de que el Châtelperroniense es un ejemplo de los neandertales imitando a la cultura de los humanos modernos. La falta de intermediarios anatómicos en estos sitios sugiere que si los neandertales encontraron y tomaron prestadas ciertas tecnologías de los Homo sapiens, no se hibridizaron.
- Un posible híbrido neandertal/sapiens de hace 24.500 años proveniente del sitio Lagar Velho, en Portugal, fue hecho público.4 Este individuo de 4 años de edad tiene un cuerpo corto y ancho como un neandertal, pero posee un cráneo anatómicamente moderno. Existe una serie de problemas en la interpretación de este hallazgo como un híbrido neandertal/sapiens.14 Primero, como un híbrido, debería tener una mezcla de caracteres en todo el cuerpo y no simplemente poseer el cuerpo de un neandertal y el cráneo de un humano moderno. Por ejemplo, si observamos a los híbridos entre leones y tigres, no poseen la cabeza de una especie y el cuerpo de la otra, sino que exhiben una mezcla morfológica de ambas especies. Segundo, y más importante, la aceptación de este espécimen como un híbrido sugeriría que los caracteres de los neandertales fueron retenidos durante entre 6.000 y 10.000 años después de que los neandertales se extinguieran, lo cual es muy poco probable. Esto es teóricamente improbable porque los caracteres de los neandertales habrían sido completamente diluidos por los genes de Homo sapiens en un periodo de tiempo tan largo.
- Los proponentes del Modelo Multirregional, tales como Milford Wolpoff,16 citan evidencia de continuidad regional proveniente de Asia. Ellos ven un enlace evolutivo directo entre los antiguos Homo sapiens [sic] de Java hasta los aborígenes de Australia. Un posible problema con esta interpretación es que los cálculos de edad de los últimos supervivientes de Homo erectus en Indonesia sugieren que vivieron allí hasta hace 50.000 años, lo cual potencialmente coincide con el tiempo de la llegada a la región de los humanos completamente modernos provenientes de África.
- La China contiene la mejor evidencia en apoyo al Modelo Multirregional. Allí se han descubierto un par de cráneos fechados en 100.000 años que parecen poseer una mezcla de caracteres del clásico Homo erectus y del Homo sapiens. Es necesario tener mejores datos de fechas geológicas y especímenes más completos para poder examinar esta posibilidad con más detalle.
Conclusión
Por el momento, la mayoría de la evidencia anatómica, arqueológica y genética da credibilidad a la perspectiva de que los humanos completamente modernos son un fenómeno evolutivo relativamente reciente. La mejor explicación que tenemos hoy en día para la génesis de los humanos modernos es el Modelo Out of Africa, el cual postula un origen singular y africano para Homo sapiens. Las innovaciones culturales y neurológicas mayores que caracterizaron a la aparición de los humanos completamente modernos resultaron ser particularmente exitosas, culminando en nuestra dominancia del planeta a costa de todas las poblaciones de los homínidos anteriores.
- Cann, R.L., M. Stoneking, and A.C. Wilson. 1987. "Mitochondrial DNA and human evolution." Nature, 325:32-36.
- Cavalli-Sforza, L.L. 2000. Genes, Peoples, and Languages. New York. North Point Press.
- Clark, G.A. and C.M. Willermet (eds.). 1997. Conceptual Issues in Modern Human Origins Research. New York. Aldine de Gruyter.
- Duarte, C, J. Maurício, P.B. Pettitt, P. Souto, E. Trinkaus, H. van der Plicht, and J. Zilhão. 1999. "The early Upper Paleolithic human skeleton from the Abrigo do Lagar Velho (Portugal) and modern human emergence in Iberia." Proc. Natl. Acad. Sci. USA, 96:7604-7609.
- Ingman, M., H. Kaessmann, S. Pääbo, and U. Gyllensten. 2000. "Mitochondrial genome variation and the origin of modern humans." Nature, 408:708-713.
- Klein, R.. 1999. The Human Career. Chicago. University of Chicago Press.
- Klein, R. 2000. "Archaeology and the evolution of human behavior." Evolutionary Anthropology, 9:17-36.
- Krings, M, H. Geisert, R.W. Schmitz, H Krainitzki, and S. Pääbo 1999. "DNA sequence of the mitochondrial hypervariable region II from the Neanderthal type specimen." Proc. Natl. Acad. Sci., 96: 5581-5585.
- Nitecki, M.H. and D.V. Nitecki (eds.). 1994. Origins of Anatomically Modern Humans. New York. Plenum Press.
- Smith, F.H. and F. Spencer (eds.). 1984. The Origins of Modern Humans: A World Survey of the Fossil Evidence. New York. Liss.
- Soffer, O. 1990. "Before Beringia: Late Pleistocene biosocial transformations and the colonization of northern Eurasia." In: "Chronostratigraphy of the Paleolithic in North, Central, East Asia and America." Novosibirisk. Acad. Of Sci. of the USSR.
- Rogers, A.R. and H.C. Harpending. 1992. "Population growth makes waves in the distribution of pairwise genetic differences." Mol. Biol. Evol., 9:552-569.
- Stringer , C. and R. McKie. 1996. Áfrican Exodus: The Origins of Modern Humanity. New York: Henry Holt.
- Tattersall, I. and J.H. Schwartz. 1999. "Hominids and hybrids: The place of Neanderthals in human evolution." Proc. Natl. Acad. Sci. USA, 96:7117-7119.
- Wolpoff, M.H. and R. Caspari. 1996. Race and Human Evolution: A Fatal Attraction. New York. Simon and Schuster.
- Wolpoff, M.H., J. Hawks, D.W. Frayer and K. Hunley. 2001. "Modern human ancestry at the peripheries: A test of the replacement theory." Science, 291:293-297.
3.2. Migraciones prehistóricas de Homo sapiens.
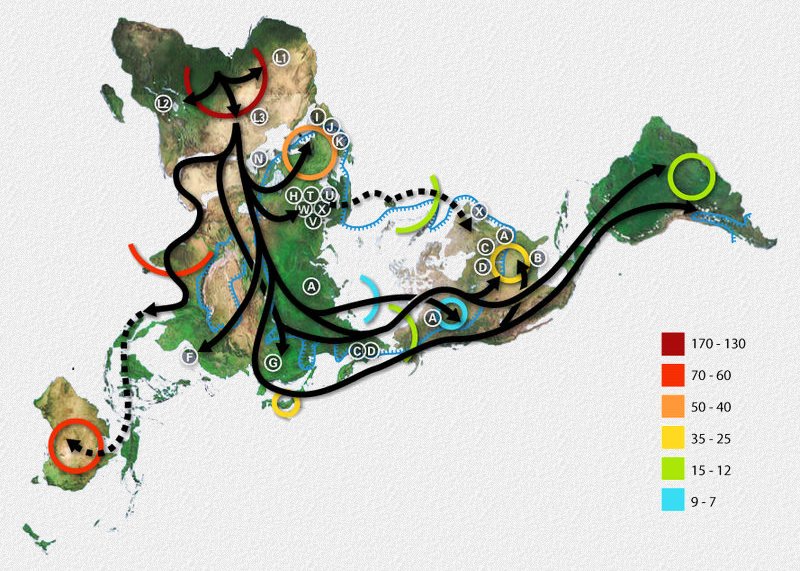
Junto a los hallazgos arqueológicos, el principal indicador de la expansión del ser humano por el planeta es el ADN mitocondrial, aunque se está investigando el cromosoma Y, que es característico de los machos, para lograr mayores precisiones.
Los humanos ya habrían comenzado a salir del África hace unos 90.000 años, colonizando para esas fechas el sur de Eurasia (estos restos fósiles han sido atribuibles a tempranos Homo sapiens, pero su real relación con los humanos modernos es aún discutida).
|
|
Mapa de la migración humana según estudios del ADN mitocondrial. La leyenda representa los miles de años desde la actualidad. La línea azul señala la extensión máxima de los hielos y las áreas de tundra durante la última glaciación.
|
3.2.1. Australia y Nueva Guinea.
Durante la última glaciación, debido a la acumulación de agua en los casquetes polares y los glaciares, el nivel del mar era unos 120 metros más bajo que el actual. Debido a esto, muchas de las islas del Sudeste Asiático se encontraban unidas al continente, permitiendo la expansión de Homo sapiens por ellas.
Sin embargo, la línea de Wallace significó para los Homo sapiens un límite insuperable durante casi 20.000 años para acceder al continente australiano, hasta que pudieron fabricar rústicas almadías o balsas de juncos para atravesar desde Indonesia el estrecho que durante las glaciaciones formaba la fosa de Wallace, separando a Australasia del continente asiático.
La línea de Wallace.
La línea de Wallace es una línea imaginaria que marca un límite biogeográfico a través de Insulindia y separa las regiones de Asia y Oceanía. La fauna, y en menor medida la flora, son notablemente distintas a cada lado, pese a la proximidad geográfica y la relativa similitud climática, reflejando historias evolutivas separadas. La línea pasa entre las islas de Bali y Lombok, al este de Java; continúa entre Borneo, que deja al oeste, y las Célebes y pasa al sur de Filipinas. Al noroeste de la línea la fauna es la característica del Sudeste Asiático; al sudeste es la australásica, que se extiende sobre Nueva Guinea, Australia y muchos archipiélagos del Pacífico sudoccidental.
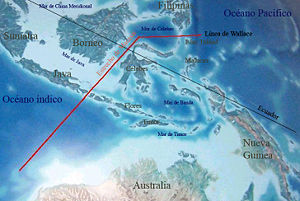
Una observación superficial y contemporánea de la línea la hace parecer imaginaria, sin embargo se corresponde con un importante límite físico, la fosa de Wallace, una extendida sima submarina producida por subducción cerca del límite entre las placas tectónicas de Australasia y de Eurasia.
Durante las fases glaciales de la glaciación cuaternaria, regiones actualmente bajo el nivel del mar a uno y otro lado de la línea quedaron al descubierto, pero separadas por un largo estrecho sobre la fosa, que resultó así una barrera para la migración de poblaciones animales. Durante el último período glacial, al oeste de la línea quedaba emergido el subcontinente de Sonda y al este el de Sahul, separados ambos por menos de 100 kilómetros. Durante la expansión de los homininos, tal fosa y tal línea resultaron un límite casi infranqueable hacia Australasia, superado inicialmente por formas modernas de Homo erectus (de las que derivó Homo floresiensis) y luego por Homo sapiens.3.2.1.1. Modelos del poblamiento de Australia.
La fecha exacta de los primeros asentamientos humanos en Australia es aún tema de debate. Sin embargo, se cree que la tierra austral ha estado habitada por seres humanos desde hace entre 42.000 y 48.000 años; en esa época hubo un período de cambio ecológico masivo que se cree fue resultado de acciones humanas. Los primeros australianos eran los ancestros de los aborígenes australianos de la actualidad; llegaron a través de puentes de tierra y pasos marítimos de poca longitud desde el sudeste asiático. La mayor parte de estas personas eran cazadores-recolectores con una compleja tradición oral y valores espirituales basados en la adoración de la tierra y en la creencia del tiempo de sueño. Los isleños del estrecho de Torres, étnicamente melanesios, habitaron desde aquel tiempo las islas del estrecho de Torres y partes del extremo norte de Queensland; poseen prácticas culturales distintas a las de los demás grupos aborígenes australianos.
Entre los cráneos humanos fósiles de Australia se han reconocido desde hace mucho tiempo dos tipologías muy diferentes. Una es, para decirlo con pocas palabras, la nuestra, que se suele calificar de grácil. La frente está levantada, la bóveda craneal es alta, no hay un verdadero torus supraorbitario, las paredes craneales no son especialmente gruesas y tampoco existen inserciones musculares particularmente marcadas en el hueso occipital o en las líneas temporales.
La otra morfología craneal es claramente más robusta, con paredes más gruesas, inserciones musculares más marcadas y un reborde óseo sobre las órbitas, pero que puede llegar a ser un auténtico torus que alcanza la región lateral.
Estas características que expresan robustez, con diferentes grados de intensidad, también se encuentran a veces en aborígenes australianos actuales. Pero, además, algunos cráneos australianos fósiles muestran una morfología general muy especial y claramente arcaica, con frentes aplanadas y bóvedas bajas, con la máxima anchura situada en posición inferior, sobre las crestas supramastoideas en lugar de en el hueso parietal, con un engrosamiento de la sutura sagital sobre la parte anterior de los parietales, con un engrosamiento de la porción final de las líneas temporales, y hasta con un torus occipital bien formado delimitado por arriba por un surco supratoral. Finalmente las caras son prognatas y los paladares, mandíbulas y dientes muy grandes.
El cráneo fósil australiano que mejor respresenta las formas robustas es el ejemplar WLH 50 de la región de los lagos del Parque Nacional de Wilandra. Este cráneo ha sido ampliamente utilizado para sostener las tesis de una continuidad evolutiva en Australasia, que se comentará a continuación. Otros fósiles australianos robustos, aunque tenidos por más recientes, son los de Kow Swamp, Cohuna y Coobool Creek. En Kow Swamp se han encontrado los enterramientos de más de 40 individuos de todas las edades, que se datan de entre hace 14.000 y 9.500 años. La muestra de Coobool Creek es también muy grande y se piensa que de edad similar la de Kow Swamp.
Los fósiles australianos de características gráciles son los de los enterramientos 1 y 3 del lago Mungo y el más moderno de Keilor, éste de hace unos 13.000 años. Los huesos del individuo 1 del lago Mungo están quemados; se trata de la más antigua cremación conocida de un cadáver. Del individuo 2 no se conserva apenas nada, mientras que el individuo 3 está completo. También se siglan como WLH 1, 2 y 3.
Estas dos morfologías craneales que se reconocen en el registro fósil de Australia admiten dos diferentes interpretaciones. Algunos autores, que se adscriben al modelo multirregional de la evolución humana, creen ver en los cráneos robustos una relación evolutiva con las poblaciones javanesas de Homo erectus. Los últimos representantes conocidos de esta especie son los cráneos de Ngandong en la terrazas del río Solo. No se conoce con precisión su edad, pero la geomorfología de la región parece indicar que las terrazas de Ngandong se depositaron en el Pleistoceno Superior, y tienen por tanto menos de 130.000 años. Recientemente se han datado estos fósiles entre 54.000 y 27.000, aunque aún deben confirmarse estas fechas con nuevos estudios, por lo que conviene considerarlas provisionales.
En su camino hacia Australia y Nueva Guinea, los Homo sapiens habrían encontrado estas poblaciones de Homo erectus y se habrían mezclado con ellas, resultando así unos australianos robustos; hay que aclarar que según este modelo evolutivo los humanos modernos y los javaneses autóctonos no serían, en realidad, especies diferentes, ya que, por definición, en ese caso no podrían producir descendientes fértiles. La coexistencia de cráneos robustos con cráneos indudablemente gráciles en Australia se explicaría invocando una doble fuente para la población de la isla. Mientras que los aborígenes robustos habrían pasado por Java, los gráciles habrían venido de China; los fósiles chinos de Liujiang y de la cueva superior de Zhoukoudian, de características modernas, y ambos del Pleistoceno Superior, podrían estar relacionados con ellos. El cráneo de Zhoukoudian sólo tiene entre 18.000 y 10.000 años, pero el de Liujiang podría pasar de los 67.000 años. Una vez en Australia, ambas poblaciones, la grácil y la robusta, se habrían cruzado en diferentes lugares y épocas.
Otro modelo muy diferente implica sustitución completa del Homo erectus por el Homo sapiens, sin mezcla alguna de sangres (del mismo modo que los neandertales habrían sido reemplazados por el Hombre de Cro-Magnon en Europa y parte de Asia). Los rasgos robustos de algunos australianos fósiles y actuales entrarían dentro de la variabilidad normal de la especie Homo sapiens. Algunos autores llegan más lejos y sostienen que las bóvedas aplanadas y las frentes huidizas son el resultado de deformaciones artificiales, una práctica cultural que se ha conocido en muchos pueblos modernos, incluidos los aborígenes australianos.
3.2.2. Europa.
Recién comenzó a ser colonizada hace sólo unos 40.000 años. Se piensa que durante milenios el desierto de Siria resultaba una barrera infranqueable desde África hacia Europa, por lo que habría resultado más practicable una migración costera desde las costas de Eritrea a las costas yemeníes y de allí al subcontinente indio. La expansión por Europa coincide con la extinción de su coetáneo de entonces, el Hombre de Neandertal, como ya se ha dicho.
3.2.3. Oceanía.
La colonización de estas islas más próximas a Eurasia se habría iniciado hace unos 50.000 años, pero la expansión por esta macro-unidad geográfica fue muy lenta y gradual, y hasta hace unos 15.000 años el Homo sapiens no comenzó una efectiva expansión por Oceanía, aunque archipiélagos como el de Hawai y Nueva Zelanda no estaban aún poblados por seres humanos hace 2.000 ó 1.500 años (se requirió el desarrollo de una apropiada técnica naval y conocimientos suficientes de náutica).
3.2.4. América.
La llegada del hombre a América o poblamiento de América es una cuestión arduamente discutida por los científicos modernos. Existe un cierto consenso, aunque no unánime, sobre el hecho de que América se habría poblado desde Siberia (Asia). Más allá de ese consenso relativo, en la primera década del siglo XXI la comunidad científica discute apasionadamente la fecha, ruta y cantidad de oleadas migratorias que poblaron el continente americano.
Se encuentra plenamente probado que durante la última glaciación, la Glaciación de Würm o Wisconsin, la concentración de hielo en los continentes hizo descender el nivel de los océanos en unos 120 metros. Este descenso hizo que en varios puntos del planeta se crearan conexiones terrestres, como por ejemplo Australia-Tasmania con Nueva Guinea; Filipinas e Indonesia; Japón y Corea.
Uno de esos lugares fue Beringia, nombre que recibe la región que comparten Asia y América, en la zona en que ambos continentes están en contacto. Debido a que el Estrecho de Bering, que separa Asia de América, tiene una profundidad de entre 30 y 50 metros, el descenso de las aguas dejó al descubierto un amplio territorio que alcanzó 1.500 kilómetros de ancho, uniendo las tierras de Siberia y Alaska, hace aproximadamente 40.000 años.
Su primera formación sucedió aproximadamente hace 40.000 años, manteniéndose unos 4.000 años. Su segunda formación se produjo hace aproximadamente 25.000 años, permaneciendo hasta hace aproximadamente 11.000-10.500, cuando volvieron a subir las aguas al final de la glaciación inundando gran parte del territorio y separando Asia de América por el Estrecho de Bering.
El dato más importante para establecer un modelo migratoria durante la última glaciación es el hecho de que Canadá estaba completamente cubierta de hielo durante la última glaciación, invadida por dos gigantescas placas: la Placa de Hielo Laurentina y la Placa de Hielo de la Cordillera. Esto hacía imposible la entrada al continente más allá de Beringia.
Apareció entonces la hipótesis del corredor libre de hielo. Según esta hipótesis, en los instantes finales de la última glaciación, comenzaron a derretirse los bordes en contacto de las dos grandes placas de hielo que cubrían Canadá, abriendo un corredor libre de hielo de unos 25 kilómetros de ancho, que seguía primero el valle del río Yukón y luego el borde este de las Montañas Rocosas por el corredor del río Mackenzie. Los científicos que sostienen la teoría estiman que esto ocurrió hace 14.000 años, aunque otros cuestionan la fecha y afirman que no pudo haber sucedido hasta hace 11.000 años, invalidando así la posibilidad que quienes originaron las culturas de Folsom y Clovis usaran esa ruta, ya que estas ya existían en esta útima fecha. Una vez abierto el corredor, los seres humanos que estaban en Beringia pudieron avanzar hacia el interior de América y dirigirse al sur. La teoría ha sido ampliamente aceptada como parte integrante del llamado Consenso Clovis, pero no hay evidencias directas que prueben el paso de seres humanos por ese corredor. Este modelo se articuló con los descubrimientos de la Cultura Clovis que databan de hace 13.500 años, para concluir que había sido integrada por los primeros migrantes que ingresaron por el Puente de Beringia, de la que a su vez habrían descendido todas las demás culturas indoamericanas.
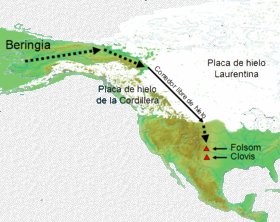
Sin embargo, cada vez existe más evidencia incontrastable de presencia humana en América anterior a los 14.000 años. Desde el punto de vista del consenso de la comunidad científica los hallazgos arqueológicos de Monte Verde (Chile), donde se ha fechado presencia humana entre hace 12.500 (Monte Verde I) y 33.000 años (Monte Verde II, aunque respecto a esta datación no hay consenso), desempeñan un papel central en la crisis del Consenso Clovis.
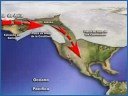
|

| Ver vídeo complementario sobre el poblamiento de América a través del corredor libre de hielo [1:35] |
Más allá de los debates en marcha y la gran cantidad de preguntas y contradicciones que se presentan en el debate científico actual es posible realizar algunas conclusiones precarias:
- Es altamente probable que el hombre americano primitivo proceda del continente asiático, especialmente de las estepas siberianas o de la región del Sudeste Asiático. Las semejanzas entre grupos poblacionales asiáticos de esas regiones y la mayoría de los aborígenes americanos ha sido objeto de análisis: etnología, lingüistica, cosmología y otros, que han permitido un enlace.
- Es probable que la dirección general de poblamiento del continente se haya producido de norte a sur. De todos modos el hecho de que las dataciones de máxima antigüedad que cuentan con consenso de la comunidad científica, Clovis (EEUU, 12.900-13.500 años) y Monte Verde (Chile, 12.500 años) se encuentren simultáneamente en América del Norte y en el extremo sur de la Patagonia, impide sacar una conclusión definitiva sobre este punto. Sin embargo estas fechas son aún muy recientes frente a otras fechas datadas en diversos lugares de América, que aún no cuentan con el consenso de la comunidad científica.
- Las culturas prehistóricas y las civilizaciones de América se desarrollaron de manera aislada al resto del planeta.
- La Revolución Neolítica americana es original y carece de toda relación con la que se produjo en la Mesopotamia asiática.
- No existen pruebas serias de la llegada a América de seres humanos después de que se cerrara el Puente de Beringia hace 11.000 años, ni existen pruebas contundentes que permitan concluir que los pueblos americanos tuvieron contactos con pueblos de otros continentes.


