2.1. Los preaustralopitecinos.
Los primeros posibles homínidos bípedos son Sahelanthropus tchadensis (con una antigüedad de 6 ó 7 millones de años), Orrorin tugenensis (unos 6 millones de años) y Ardipithecus (entre 5,5 y 4,5 millones de años). Debido a la escasez de restos fósiles y a lo fragmentario de los mismos, no hay acuerdo general sobre si estos primeros homínidos fueron totalmente bípedos.
2.1.1. Sahelanthropus tchadensis.
Sahelanthropus tchadensis es una especie de homínido fósil hallada en el desierto del Djurab por un equipo franco-tchadiense. El único espécimen, apodado Toumaï (en la imagen), se ha datado en 6 a 7 millones de años de antigüedad.

Su descubrimiento se realizó el 19 de julio de 2001 por el equipo dirigido por Alain Beauvilain en la región de Toros Menalla de la actual República de Chad.
Se cree que vivió en zonas pantanosas. Su cráneo es simiesco, aunque de rostro breve y dientes pequeños, en particular los colmillos, lo cual le acerca a los humanos.
Se han encontrado un cráneo, dos fragmentos de mandíbula inferior y tres dientes aislados. No presenta cresta craneal; su volumen cerebral es de 350 cm3. Tiene rostro alto y poco prognato, caninos relativamente pequeños, y ausencia de espacio entre los dientes (diastema). Podría ser el antecesor de Ardipithecus ramidus.
2.1.2. Orrorin tugenensis.
Orrorin tugenensis es una especie de homínido fósil encontrado en las proximidades de la localidad de Tugen, en el área montañosa central de la actual Kenia, por la paleoantropóloga francesa Brigitte Senut y el inglés Martin Pickford.
Como el descubrimiento se dio a conocer en 2001, en un principio se habló erróneamente de Homo milenium, y aún se mantiene la denominación paralela Milenium ancestor para referirse al Orrorin tugenensis. Sin embargo, para la comunidad antropológica resultó patente desde un primer momento que esta especie no correspondía al género Homo, ni siquiera al conjunto de los australopitecinos. La datación demostró que los Orrorin tugenensis vivieron hace al menos unos 6 millones de años.
La morfología de los Orrorin era notablemente similar a la de los actuales chimpancés con una importante diferencia: la longitud y forma del húmero y del fémur así como la disposición de la articulación con la pelvis evidencian que estos homínidos estaban capacitados para la bipedestación. La talla de los ejemplares hallados es de 1,4 metros.
Otra característica morfológica muy interesante es su dentadura: con caninos pequeños y molares bastante grandes, se deduce que estos primates tenían una dieta principalmente herbívora y frugívora, aunque es muy probable que fueran omnívoros y obtuvieran sus proteínas alimentándose también de insectos.
El género Orrorin es, posiblemente, uno de los eslabones que ha derivado en el ser humano actual, y se considera también posible que sea descendiente directo del Sahelanthropus tchadensis y ancestro directo del Ardipithecus.
2.1.3. Ardipithecus.
El género Ardipithecus comprende a un conjunto de primates homínidos fósiles hallados en Etiopía a los que se considera descendientes de los Orrorin tugenensis y directos ancestros de los australopitecinos. Las similitudes con los Australopithecus son grandes, aunque los fósiles encontrados presentan rasgos más simiescos y menos corpulencia que éstos. Esto ha llevado a algunos investigadores a afirmar que se encuentra en la línea evolutiva del chimpancé, pero la mayoría considera, basándose en la similitud de su dentadura con la de los Australopithecus, que se encuentran insertos en la línea evolutiva humana.
En la actualidad se han clasificado dos especies de Ardipithecus: el Ardipithecus ramidus ("ramid" significa "raíz" en la lengua afar de Etiopía y hace referencia a la proximidad de esta especie a las raíces de la humanidad) y el Ardipithecus kadabba, aunque este último se considera a veces una subespecie del primero con el nombre de Ardipithecus ramidus kadabba. Los fósiles de Ardipithecus ramidus están datados en un periodo de entre 4,5 y 4,1 millones de años, mientras que se considera que Ardipithecus kadabba tiene una antigüedad de entre 5,5 y 5,8 millones de años.
Basándose en el tamaño de los huesos, se cree que el género Ardipithecus tenía la envergadura de un chimpancé actual. La forma de los dedos de los pies del Ardipithecus ramidus sugiere que andaban erguidos, y esto plantea un problema con las teorías actuales sobre los orígenes de la bipedestación.
Ardipithecus ramidus se distingue como homínido de los grandes simios actuales y algunos restos de hominoideos fósiles en:
- Morfología del canino más incisiviforme (con forma de incisivo). En todos los homininos los caninos presentan dos pequeñas cúspides laterales accesorias a la principal, rompiendo la forma cónica típica de los caninos primitivos.
- Patrón de desgaste plano en el canino inferior. El desgaste de los caninos en los grandes monos (lo primitivo) es muy vertical (como una cizalla) y forma bordes cortantes por delante y por detrás. En los homininos (incluido Ardipithecus ramidus) esta función se pierde y el desgaste es plano.
- Foramen magnum localizado anteriormente con respecto al foramen carotideo (orificio por el que entran las arterias carótidas en el cráneo). El foramen magnum se sitúa en una posición aparentemente más vertical que en los simios. Con restos tan fragmentarios, esta evidencia resulta poco concluyente para determinar si Ardipithecus ramidus era bípedo o no lo era.
En cambio, Ardipithecus ramidus se distingue de otras especies de homininos, incluyendo a los primeros australopitecinos, en:
- Caninos grandes en relación con la dentición postcanina (premolares y molares). Más grandes que los de cualquier hominino pero menos grandes que los de un chimpancé.
- Primer molar inferior deciduo (primera muela de leche) estrecho y oblicuamente alargado, con una sola cúspide principal, lo que corresponde a una morfología muy primitiva, más próxima a la de los chimpancés actuales.
- Caninos y molares con esmalte dental fino. El resto de los homininos presenta el esmalte dental grueso. El espesor del esmalte de los dientes parece estar fuertemente relacionado con el tipo de dieta. A un esmalte fino, como el de Ardipithecus ramidus, los chimpancés y demás simios actuales, se le atribuye una funcionalidad en la ingestión de frutas y bayas (dieta blanda). El esmalte grueso que presentan los demás homininos conocidos se relaciona con un cambio hacia una dieta más abrasiva.
- Tercer premolar inferior y superior más pronunciadamente asimétrico, prácticamente con una sola cúspide. La hominización de los dientes se supone que está relacionada con un aumento en el número y tamaño de las cúspides especialmente en los premolares. Se conoce como molarización. En esta característica Ardipithecus ramidus también es primitivo.
2.2. Los australopitecinos.
Los primeros homínidos de los que se tiene la seguridad de que fueron completamente bípedos son los miembros del género Australopithecus, de los que se han conservado esqueletos muy completos (como el de la famosa Lucy).
Este tipo de hominino prosperó en las sabanas arboladas del este de África entre 4 y 2,5 millones de años atrás con notable éxito ecológico, como lo demuestra la radiación que experimentó, con al menos cinco especies diferentes esparcidas desde Etiopía y el Chad hasta Sudáfrica.
Su desaparición se ha atribuido a la crisis climática que se inició hace unos 2,8 millones de años y que condujo a una desertificación de la sabana, con la consiguiente expansión de los ecosistemas abiertos, esteparios. Como resultado de esta presión evolutiva algunos Australopithecus se especializaron en la explotación de productos vegetales duros y de escaso valor nutritivo, desarrollando un impresionante aparato masticador, originando el género Paranthropus; otros Australopithecus se hicieron paulatinamente más carnívoros, originando a los primeros Homo.
Las características principales del género Australopithecus son:
- En el cráneo:
- Capacidad craneal relativamente pequeña.
- Paredes craneales delgadas.
- Bordes supraorbitarios robustos.
- Protuberancia occipital externa por debajo del plano de Frankfurt.
- Flexión basicraneal.
- Posición anterior de los cóndilos occipitales.
- Orientación del foramen magnum en un plano casi horizontal.
- Prognatismo subnasal moderado.
- Huesos maxilares masivos.
- En la mandíbula:
- Mandíbulas masivas y robustas.
- Arco mandibular interno en "V" o "U" aguda.
- Arcada dental parabólica sin diastema.
- En los dientes:
- Caninos espatulados con desgaste perpendicular al diente.
- Premolares y molares absoluta y relativamente grandes.
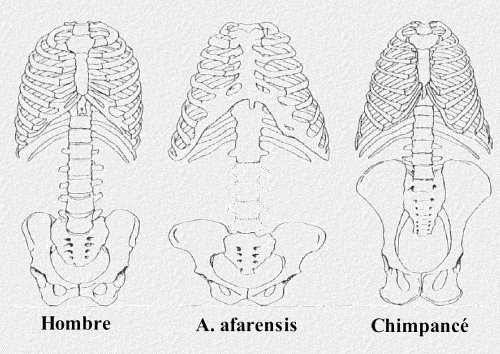
La anatomía de los australopitecinos muestra asimismo una serie de adaptaciones para la bipedestación, tanto en la parte superior como inferior del cuerpo. Entre las adaptaciones de la parte inferior se incluyen las siguientes:
- El ilion era mucho más corto y ancho que en los simios, lo que permitía a los músculos equilibrar el cuerpo tras cada paso.
- La pelvis tenía forma cóncava para alojar los órganos internos durante la postura erguida.
- La parte alta de los miembros inferiores formaban un ángulo hacia el interior desde la articulación de la cadera, permitiendo así a las rodillas soportar mejor el peso del cuerpo al andar erguido. Por el contrario, los miembros inferiores de los simios están colocados casi en sentido vertical desde la cadera, de forma que cuando andan erguidos su cuerpo se balancea hacia los lados.
- Los dedos de los pies eran más cortos y menos flexibles que los de los simios, de forma que actuaban como palancas para impulsar el cuerpo a cada paso.
Por encima de la pelvis también se produjeron otras adaptaciones. La columna australopitecina presentaba una curva en "S" que disminuía la longitud total del torso y le confería rigidez y equilibrio cuando se encontraba erguido; los simios, por el contrario, tienen una columna relativamente recta. El cráneo australopitecino también presentaba una adaptación importante relacionada con la bipedestación: el foramen magnum se encontraba en una posición más adelantada que en los simios, lo que permitía a la cabeza mantenerse en equilibrio sobre la columna erguida.
Está claro que los australopitecinos caminaban erguidos sobre el suelo, pero los paleoantropólogos no tienen una opinión unánime sobre si pasaban también una parte importante de su tiempo en los árboles. Algunas características físicas como, por ejemplo, los dedos curvos y largos y los brazos alargados, parecen confirmar que así era. Sin embargo, los dedos, a diferencia de los de los simios, tal vez no eran lo suficientemente largos como para permitirles balancearse de rama en rama.
2.2.1. Australopithecus anamensis.
Australopithecus anamensis es una especie de hominino de 4,2-3,9 millones de años de antigüedad encontrada en Kenia y descrita en 1995 por Meave Leakey. El nombre de esta especie proviene de la palabra tukana "anam" que significa lago y fue elegida en razón de la proximidad de Kanapoi al lago Turkana, y como recordatorio del gran lago que existió en estas desiertas zonas en esa época, el lago Lonyumun.
Sus muelas poseían gruesos esmaltes, por lo que se deduce que no comía sólo hojas y frutos, sino alimentos más duros. Vivió en un ambiente forestal, pero más abierto que el de sus supuestos antepasados los Ardipithecus, y se sabe que sí caminaba erguido. Presentaban un claro dimorfismo sexual en tamaño corporal. Es posible que formaran comunidades de varios machos emparentados, en las que cada uno agruparía un pequeño harén de hembras.
El equipo de Tim White encontró en diciembre de 2005 restos de esta especie en el sitio Asa Issie, al noreste de Etiopía, en el valle medio del río Awash. Datados con entre 4,1 y 4,2 millones de años de antigüedad, son un fémur, fragmentos de maxilar y algunos dientes, incluyendo el canino más grande hallado entre los homininos. Habían sido encontrados además otros 9 fósiles de esta especie, en Kanapoi, Kenia, la mayoría en 1994; más otros 12 fósiles, principalmente dientes, encontrados en 1988, en Allia Bay, Kenia y un fragmento de húmero que había sido encontrado en 1965 por Bryan Patterson y fue identificado posteriormente.
Aunque los dientes y una mandíbula presentan aún similitud con los simios, tienen, por primera vez en la evolución de los homininos, un esmalte grueso. El fragmento de húmero es semejante al de los humanos y la parte de tibia encontrada en 1994 por Kamoya Kimeu y el fémur de Asa Issie son la más antigua evidencia del bipedismo erecto de los homininos. A partir de las características anatómicas de los huesos de los brazos (largos brazos y las falanges curvadas) y de las piernas se deduce que Australopithecus anamensis era, al mismo tiempo, bípedo y un poderoso trepador. Su hábitat, en comparación al de los homínidos que lo preceden, era un lugar más ideal para la evolución del bipedismo, con regiones menos arboladas y sabanas.
2.2.2. Australopithecus afarensis.
Australopithecus afarensis fue un hominino que vivió entre los 3,9 y 3 millones de años antes del presente. Era de contextura delgada y grácil, y se cree que habitó sólo en África Oriental (Etiopía, Tanzania y Kenia). Es aceptado entre la mayoría de la comunidad científica que puede ser uno de los ancestros del género Homo.

Fue descubierto el 24 de noviembre de 1974 por Donald Johanson, Yves Coppens y Tim White en Hadar, Etiopía. El espécimen encontrado en aquel momento fue el mundialmente conocido como Lucy (en la imagen).
Este descubrimiento destacó sobre los demás por muchas cualidades, especialmente por ser el Australopithecus mejor conservado a través de tantos años. Así fue posible comprobar que su capacidad para caminar erguido, como los humanos actuales, precedió al crecimiento del cerebro. El estudio de su dentadura aclaró aspectos fundamentales sobre la evolución de los homininos y descubrió la evolución simultánea de géneros, de manera que la línea Paranthropus se apartó de otras y en particular de la que evolucionó hacia Homo.
Los restos de Lucy fueron encontrados en el lugar donde habitaba la tribu Afar, de ahí el nombre afarensis, y junto con otros doce individuos de la misma especie, incluido cráneos, conforman lo que se llama "la primera familia".
Australopithecus afarensis es, como todo Australopithecus, un primate bípedo erguido, pero diferente a nosotros en varios aspectos:
- Capacidad craneal bastante menor: 375 a 550 cm3. Esto significa un cerebro de un tamaño cercano a la tercera parte del humano actual promedio.
- Australopithecus afarensis era un hominino con frecuente actividad arbórea, ya sea recolectando frutos, durmiendo, jugando, etc.
- Su pecho no era en forma de barril, como el nuestro, sino que se estrechaba agudamente hacia arriba (forma de campana).
- El dimorfismo sexual era muy marcado, con los machos mucho más corpulentos que las hembras.
En lo que se refiere a sus sistema óseo es muy importante recalcar que tenían el cráneo relativamente pequeño, como los antropomorfos, aunque en comparación con el tamaño del cuerpo era relativamente grande. Además, su cara era bastante grande y se proyectaba delante del cráneo (prognatismo), debido al tamaño de los dientes.

A diferencia de los antropomorfos, los caninos de Australopithecus afarensis son reducidos aunque se proyectan ligeramente delante del diente adyacente. Los incisivos son grandes (asociados al régimen frugívoro). Los molares y premolares son de tamaño sustancial, con superficies planas.
De su boca también se concluye que el paladar es muy similar al del hombre actual porque, aunque grande, forma una curva que no es parabólica, ni de lados paralelos, como en los grandes simios.
La forma de la pelvis es importante ya que de ella se han realizado estudios necesarios para determinar el caminar bípedo erguido de los Australopithecus; es de pequeño tamaño en comparación a la de los antropomorfos y presenta un canal de parto, en la hembra, más pequeño que el actual y con una notoria forma de riñón. Las hembras de la especie no necesitaban tener un gran canal de parto, debido al pequeño tamaño del cráneo de las crías.
 Las pisadas de Laetoli.
Las pisadas de Laetoli.

Las icnitas de Laetoli se formaron sobre cenizas muy finas expulsadas por el volcán Sadimán hace 3,6 millones de años. Tras la erupción llovió (hay marcas de gotas de lluvia) y el suelo se humedeció. Los homínidos y otros animales pasaron y dejaron su rastro. El sol salió y secó rápidamente los rastros y, dado el alto contenido en carbonatos, se endureció como el cemento. Cayeron otras capas de cenizas hasta cubrir unos 16-20 centímetros y las huellas quedaron enterradas y conservadas.

Presentan un patrón morfológico totalmente humano. El golpe de talón está muy marcado, el dedo gordo está completamente alineado al resto de los dedos y el arco plantar medial está bien desarrollado. Las relaciones espaciales entre las huellas presentan un patrón sorprendentemente humano (distancia entre las pisadas, orientación de las mismas, etc.).
Se distinguen tres rastros de pisadas. En los años setenta del siglo XX se descubrieron rastros de dos individuos desplazándose hacia el norte. Son dos rastros paralelos, separados entre sí 25 centímetros, demasiado próximos para dos personas caminando una al lado de la otra. Es posible que siguieran el mismo camino pero que no se produjeran de forma simultánea, dado que las condiciones de ambos rastros son algo diferentes. El primero de los rastros (conocido como rastro G-1), el de pies más pequeños, presenta los contornos nítidos, como si se hubieran formado sobre una superficie firme y compacta (húmeda), mientras el rastro del pie mayor (rastro G-2) tiene los contornos más difusos y agrandados, como si se hubieran producido más bien sobre una superficie seca y polvorienta. El rastro G-1 parece detenerse a mitad de camino, dar media vuelta hacia la izquierda, y volver a retomar el camino inicial.
No fue hasta después cuando los investigadores se dieron cuenta de que el rastro situado más al este (rastro 2) representaba huellas superpuestas de dos individuos: G-2 la más profunda y G-3 la más superficial.
A partir de las huellas se han hecho estimaciones de la estatura, pero con varios presupuestos de partida:
- Los homininos de Laetoli tienen las mismas proporciones del pie que los humanos modernos.
- Los individuos representados son adultos.
- Las pisadas son buenos indicadores del tamaño del pie.
Usando varias poblaciones actuales, el rango obtenido para la mayor de las pisadas es de 1,34 a 1,56 metros y para la de menor tamaño de 1,15 a 1,34 metros. estas estimaciones son consistentes con las obtenidas a partir de restos del esqueleto postcraneal de Australopithecus afarensis, el candidato más adecuado como responsable de estas pisadas.
Este rastro permite estudiar el tipo de locomoción de los homininos que lo produjeron. En las huellas de Laetoli, como en las de un humano actual, la transmisión del peso empieza en el talón, sigue a lo largo del borde exterior y termina en el pulgar, como cabría esperar en un bípedo.
En este yacimiento (Laetoli) se han encontrado restos fósiles de Australopithecus afarensis, lo que permite especular con bastante confianza que individuos de esta especie son los autores de las huellas.

|

| Ver vídeo complementario sobre Australopithecus afarensis y las icnitas de Laetoli [5:17] |
2.2.3. Australopithecus bahrelghazali.
Australopithecus bahrelghazali es una especie fósil de hominino hallada en 1995 por Michel Brunet en Bahr el-Ghazal, Chad, representado por una mandíbula con siete dientes de 3,5 a 3 millones de años de antigüedad. Fue apodado Abel. Es la primera evidencia de fósiles de homininos al oeste del valle del Rift. Se considera que representa una línea de Australopithecus distinta de la que evolucionó hacia Homo.
East Side Story.
East Side Story es una hipótesis propuesta por Yves Coppens en 1994 que intenta explicar la aparición de los homínidos bípedos.
Esta hipótesis se basa en los siguientes hechos:
- La zona oeste de África comprendida entre el océano Atlántico y el valle del Rift está poblada por ecosistemas selváticos, mientras que en la zona al este del valle del Rift hasta el océano Índico predominan los ecosistemas de sabana.
- Los homínidos bípedos más antiguos se han hallado al este del valle del Rift.
Según el mencionado autor, durante el Mioceno, un gran cinturón de selva se extendía desde el Atlántico hasta el Índico. La formación del valle del Rift (finales del Mioceno) debido a procesos tectónicos, con los consiguientes cambios en el relieve, levantó grandes barreras montañosas que han actuado desde entonces como pantalla orográfica; así los vientos cargados de humedad provenientes del Atlántico no pueden superar tal pantalla y al este de la misma se reduce la precipitación. Los grandes simios que quedaron aislados en poblaciones occidentales (antepasados de gorilas y chimpancés) y poblaciones orientales. Éstas debieron adaptarse al nuevo ecosistema más seco (sabana, bosque abierto) lo que las empujó a abandonar la vida arborícola para hacerse habitantes del suelo. La mejor solución para un homínido que se ve obligado a desplazarse en el suelo para buscar alimento y refugio en árboles vecinos sería el bipedismo.
La hipótesis ha sido criticada tras el hallazgo de los fósiles de Australopithecus bahrelghazali en el Chad, al oeste del valle del Rift, así como de restos de homínidos supuestamente bípedos no asociados a ecosistemas abiertos (Orrorin tugenensis, Sahelanthropus tchadensis, Ardipithecus). El Gran Valle del Rift.
El Gran Valle del Rift es una gran fractura geológica cuya extensión total es de 4.830 kilómetros en dirección norte-sur. Aunque generalmente se habla de este valle para referirse sólo a su parte africana, desde Yibuti a Mozambique, lo cierto es que el Mar Rojo y el valle del Jordán también forman parte de él. Comenzó a formarse en el sureste de África (donde es más ancho) hace unos 30 millones de años y sigue creciendo en la actualidad, tanto en anchura como en longitud, expansión que con el tiempo se convertirá en una dorsal oceánica (de hecho, ya lo es en la zona del Mar Rojo gracias a su comunicación con el Océano Índico). Los constantes temblores de tierra y emersiones de lava contribuyen a este crecimiento y, de seguir a este ritmo, el fondo del valle quedará inundado por las aguas marinas de forma total dentro de 10 millones de años. Con ello, África se habrá desgajado en dos continentes distintos que procederán a separarse más aún hasta formar un nuevo océano.
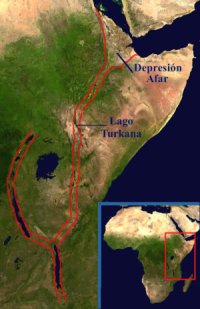
A la altura de África Central se divide en dos valles distintos que vuelven a unirse más al sur, en Tanzania. El del este acoge en su interior grandes extensiones de sabana por las que se mueven enormes manadas de mamíferos, como el búfalo africano, el ñu, la cebra de planicie, la jirafa o el elefante africano. En la del oeste, en cambio, predomina la selva y allí pueden verse chimpancés y gorilas, entre otros animales. El sistema del Rift también acoge a la mayor elevación del continente africano, el volcán Kilimanjaro y a algunos de los mayores lagos africanos, como el Turkana, el Tanganica o el Malawi. También se considera al lago Victoria, el segundo lago más grande del mundo, como parte del sistema, aunque en realidad se encuentra en el terreno situado entre los dos ramales antes citados. Entre las cordilleras que corta o siguen a lo largo del valle están el macizo etíope y los montes Mitumba. También son abundantes los lagos salados, tanto presentes como ya secos, que emergen a través de las grietas en el terreno y no son producto de la lluvia.
La rotura del continente africano ejercida por el Gran Valle del Rift es la responsable también de que en el este del continente el clima sea más seco que en el oeste. Debido a ello, en esta parte de África apareció primero la sabana, y los simios locales, que hasta entonces vivían en las ramas de los árboles, debieron hacerse terrestres y dieron lugar a los primeros homínidos. La gran grieta ha dejado también al descubierto cientos de metros de estratos geológicos, por lo que los fósiles y la historia geológica en general de esta parte de África son los mejor conocidos de todo el continente negro.2.2.4. Australopithecus garhi.
Australopithecus garhi es una especie de hominino cuyos fósiles se descubrieron en 1996 por un grupo de investigadores liderado por el paleontólogo etíope Berhane Asfaw y el antropólogo estadounidense Tim White.
Inicialmente se creyó que era el eslabón perdido entre los géneros Australopithecus y Homo, y por lo tanto un ancestro de nuestra especie. Sin embargo Australopithecus garhi es más avanzado que cualquier otro australopiteco, y una especie contemporánea (o casi) de las especies ancestrales de Homo, por lo tanto no es probable que sea un ancestro humano. Los restos hallados proceden de un lapso temporal con un escaso registro fósil, entre 2 y 3 millones de años. Tim White fue el científico que halló el primero de los fósiles de Australopithecus garhi en 1996 cerca de la ciudad de Bouri, en el río Awash medio de la Depresión de Afar, en Etiopía. La especie fue confirmada y establecida como Australopithecus garhi el 20 de noviembre de 1997 por el paleoantropólogo etíope Yohannes Haile-Selassie. El nombre de la especie "garhi" significa "sorpresa" en la lengua local Afar.
Los rasgos de Australopithecus garhi son algo distintos de los típicamente vistos en Australopithecus afarensis y en Australopithecus africanus. Un ejemplo de las diferencias puede verse al comparar el maxilar encontrado en Hadar, Etiopía (Australopithecus afarensis) con el espécimen de Bouri de Australopithecus garhi. La capacidad craneal de Australopithecus garhi es de 450 cm3, parecida a la de otros australopitecos. La mandíbula hallada por Asfaw tiene una morfología general compatible con la misma especie, aunque es posible que pertenezca a otras especies de homininos encontradas en los mismos depósitos. Los estudios hechos en los premolares y molares muestran cierta similitud con los de Paranthropus boisei, que son más grandes que en otras formas gráciles de australopitecos. Se ha sugerido que si Australopithecus garhi es un ancestro de Homo la morfología maxilar debería haber tenido que evolucionar rápidamente, en apenas 200.000 ó 300.000 años.
Los pocos artefactos primitivos de piedra descubiertos con los fósiles de Australopithecus garhi, datados en 2,5-2,6 millones de años, recuerdan vagamente a la tecnología Olduvayense. Estas herramientas son más antiguas que las de Homo habilis, que es considerado un posible ascendiente directo de homininos más modernos. Asimismo, en Bouri se encontraron unos 3.000 artefactos toscos de piedra de una edad estimada en 2,5 millones de años. Estas antiguas herramientas (cantos tallados y lascas) muestran el uso de varias técnicas que pueden observarse en las industrias más tardías Olduvayense y Achelense.
Junto con los fósiles homininos se han encontrado también huesos de antílopes los cuales muestran marcas de corte curvilíneas realizadas por instrumentos de piedra. Algunos de los huesos de antílope de los miembros muestran evidencias de haber sido abiertos a golpes con martillos de piedra. Esto indicaría que estos homininos eran capaces de adquirir grandes cantidades de recursos alimenticios de alta calidad (carne y grasa de la médula ósea).
Desde el punto de vista de la tecnología y de la dieta, Australopithecus garhi representa una primera etapa hacia el surgimiento del género Homo. Antes del descubrimiento de Australopithecus garhi, el antecedente más antiguo de un hominino capaz de utilizar un instrumento de piedra estaba representado por el género Homo, con una antigüedad de 2,3 millones de años y encontrado también en la misma región. La relación entre Australopithecus africanus y el género Homo era muy difícil de descifrar debido al vacío existente en el registro fósil del África del este entre los 2 y 3 millones de años. Australopithecus garhi viene a llenar este vacío y a demostrar que probablemente el uso de instrumentos y el cambio de dieta se produjo antes del aumento de la capacidad craneal.
2.2.5. Australopithecus africanus.
Australopithecus africanus es una especie de hominino fósil de Sudáfrica, datado en unos 5 millones de años. La historia de Australopithecus africanus como especie reconocida data de 1924, cuando fue descubierto por Raymond Dart en Taung, Sudáfrica, el cráneo de un niño conocido como el Niño de Taung. Pero no fue tomado en cuenta hasta 30 años después.
De acuerdo con los estudios dentales realizados, Australopithecus africanus procede de Australopithecus afarensis y hasta no hace mucho se creía que abría una línea evolutiva hacia el género Paranthropus, en tanto que otra línea, que también procede de Australopithecus afarensis, estuvo representada por Homo habilis y culminó en el Homo sapiens.
Entre las características se resalta la forma de caminar bípeda, común a todos los homininos. Tenían actividad arbórea en forma activa. Su peso promedio oscilaba entre los 33 y 67 kilogramos, con una estatura de 1,50 metros de alto.
Sus características óseas son de gran importancia para diferenciarse del resto de Australopithecus: su capacidad craneana es de 480 a 520 cm3. Su caja craneal es más alta y redondeada que la de los antropomorfos, y su esqueleto postcraneal similar al del Australopithecus afarensis.

Su cara, desde un punto de vista general, es más corta. Presenta menor prognatismo (la mandíbula está adelantada con respecto al maxilar), unido a un menor tamaño de piezas dentales. Éstas no se diferencian mucho de las del humano. Los caninos eran pequeños, sin sobresalir como en los antropomorfos, y los incisivos eran tan pequeños como los nuestros. También es importante mencionar que desaparece el diastema (separación entre incisivos), o es muy rara su existencia. Por tanto, notamos una reducción de caninos e incisivos y hay un mayor énfasis de la masticación en el resto de la dentadura.
2.2.6. Kenyanthropus platyops.
Kenyanthropus platyops es el nombre propuesto para una especie de hominino de la que se encontró un cráneo fósil en 1999. El hallazgo fue hecho por un equipo dirigido por Meave Leakey y su hija Louise, en Kenia, en una región semidesértica cercana al lago Turkana. El fósil data de unos 3,5 millones de años. Tiene características muy interesantes, ya que posee rasgos faciales planos y delicados, y dientes relativamente pequeños.
Los científicos han propuesto definir el género Kenyanthropus, pues el cráneo encontrado muestra una mezcla equilibrada de rasgos de Australopithecus y Homo, se parece al "hombre 1470", otro fósil hallado hace unos años en esa región, clasificado como Homo rudolfensis, y con una datación mucho menor, de unos 1,8 millones de años. No obstante, dado que el espécimen tipo (KNM-WT 4000) está terriblemente distorsionado, hay autores que no lo consideran ni un género ni una especie válida y que podría tratarse simplemente de un espécimen de Australopithecus afarensis o incluso de alguna especie de Homo, tal como Homo rudolfensis.
2.2.7. Paranthropus.
Paranthropus es un género extinto de homínidos bípedos, del que se han descrito tres especies, caracterizado por una gran robustez de la mandíbula y los molares. Los Paranthropus (parántropos o australopitecinos robustos) descienden probablemente del género Australopithecus (australopitecinos gráciles). Algunos paleontólogos consideran, sin embargo, que los australopitecinos robustos pertenecen al género Australopithecus.
Los fósiles encontrados corresponden a especies que vivieron entre 2,6 y 1,1 millones de años antes del presente. Se caracterizan por un aparato masticador especializado, constituido por grandes mandíbulas y molares con capa de esmalte muy gruesa, unos incisivos y caninos muy pequeños y unos premolares que se desarrollan como si de molares se tratase, y músculos faciales poderosos que se insertaban en una cresta sagital similar a la del gorila. Sus cerebros tenían entre 410 cm3 los más tempranos y 530 cm3 el resto, siendo por tanto inferiores a los del género Homo.
Hasta mediados de los años ochenta del siglo XX se aceptaba que el grupo de los parántropos se había originado a partir de un hominino muy próximo a Australopithecus africanus, si no de esta misma especie. Según este punto de vista, la especie Paranthropus robustus sería la más primitiva y a partir de ella habría aparecido la variante esteafricana: Paranthropus boisei. Sin embargo, a medida que se iban encontrando fósiles cada vez más antiguos de Paranthropus boisei, se hacía cada vez más difícil de sostener este punto de vista, ya que la especie pretendidamente descendiente (Paranthropus boisei) parecía más antigua que la presunta antepasada (Paranthropus robustus). Para conciliar esta aparente contradicción, se argumentaba que los fósiles más antiguos de Paranthropus robustus aún no habían sido descubiertos.
Sin embargo, la respuesta a este acertijo había sido ya encontrada a muchos miles de kilómetros de allí, aunque casi nadie se había percatado de ello. En 1967, dos científicos franceses, Camille Arambourg e Yves Coppens, dieron a conocer una mandíbula de hominino hallada en Etiopía en uno de los yacimientos del curso bajo del río Omo, cerca de su desembocadura en el lago Turkana, y conocida como Omo 18-1967-18. La antigüedad del fósil está establecida en cerca de 2,6 millones de años y fue atribuido por su descubridores a una especie nueva: Paraustralopithecus aethiopicus.
Casi veinte años después, en 1986, Richard Leakey y sus colaboradores publicaron el hallazgo de un extraordinario cráneo fósil procedente de uno de los yacimientos de la ribera occidental del lago Turkana en Kenia. Este fósil, de una antigüedad cercana a los 2,5 millones de años, recibe oficialmente el nombre de KNM-WT 17000 (habitualmente abreviado a WT 17000), aunque también se le conoce más familiarmente como el Black Skull (Cráneo Negro), debido al color del manganeso que impregnó el hueso durante el proceso de fosilización. Los autores del hallazgo propusieron que el nuevo cráneo pertenecía a la misma especie que la mandíbula hallada por Arambourg y Coppens dos décadas antes y, aunque se mostraron partidarios de incluir ambos restos dentro de una forma primitiva de la especie Australopithecus boisei, no descartaron que pudieran pertenecer a otra especie cuyo nombre debería ser Australopithecus aethiopicus. Esta última posibilidad ha sido la que ha ido ganando más partidarios en los últimos años (bien como Australopithecus aethiopicus o bien como Paranthropus aethiopicus). Los fósiles más modernos atribuidos a Paranthropus aethiopicus son una serie de restos craneales y mandibulares procedentes de los yacimientos del río Omo, fechados entre 2,3 y 2,1 millones de años de antigüedad.
Hoy conocemos lo suficiente de la anatomía de Paranthropus aethiopicus como para establecer que es más primitiva que la de sus congéneres Paranthropus robustus y Paranthropus boisei. Muchos de los rasgos que caracterizan la anatomía de los parántropos más modernos aún no están presentes en Paranthropus aethiopicus, y otros sólo lo están de modo incipiente. Para la práctica totalidad de los paleoantropólogos no hay duda: Paranthropus aethiopicus es el perfecto intermedio evolutivo entre Australopithecus afarensis, por una parte, y Paranthropus robustus y Paranthropus boisei, por la otra, lo que descarta a Australopithecus africanus, o una forma similar, como antepasado directo de los parántropos.
En cuanto a las relaciones evolutivas entre Paranthropus robustus y Paranthropus boisei, la mayor parte de los investigadores coinciden en aceptar que ambas derivan independientemente de Paranthropus aethiopicus, si bien Paranthropus boisei sufrió una especialización más acentuada y Paranthropus robustus retuvo más caracteres primitivos.
2.2.7.1. Paranthropus aethiopicus.
Paranthropus aethiopicus es un hominino fósil que vivió África Oriental hace entre 2,6 y 2,2 millones de años. Tenía poderosos músculos masticadores que se sujetaban al cráneo gracias a una prominente cresta sagital similar a la de los gorilas. Esta cresta es imprescindible para que la enorme musculatura facial pueda sujetarse en el cráneo. Tenía una dieta estrictamente vegetariana a base de vegetales duros. Debió vivir en un medio ambiente más seco que los Australopithecus, aunque parece que fuera un ambiente pleno de sabana.

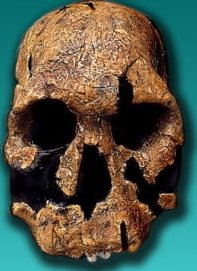
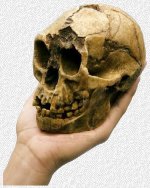
En 1967 se encontró cerca del lago Turkana, en la zona etíope, una mandíbula de unos 2,5 millones de años de antigüedad. El descubrimiento no tuvo continuidad hasta que en 1985 el paleoantropólogo estadounidense Alan Walker descubrió el antes mencionado Black Skull (en la imagen), perteneciente a la misma especie que la mandíbula hallada en 1967.
Este fósil hizo posible la identificación de una nueva especie que representaba el eslabón entre Australopithecus afarensis y Paranthropus boisei. Esta especie intermedia presenta rasgos propios de los Australopithecus y anuncia los que van a caracterizar tanto al Paranthropus boisei como al Paranthropus robustus.
2.2.7.2. Paranthropus boisei.
Paranthropus boisei es una especie de hominino fósil conocida de África Oriental, que vivió en un entorno seco y se alimentaba de vegetales duros, para lo que desarrolló un imponente aparato masticador destinado a triturar semillas y raíces. Aparece en el registro fósil hace 2 millones y desaparece hace un millón de años.
El primer ejemplar conocido fue un cráneo descubierto por Mary Leakey el 17 de julio de 1959 en la garganta de Olduvai, Tanzania; vivió hace 1,75 millones de años. Mary y su esposo Louis Leakey clasificaron inicialmente la especie como Zinjanthropus boisei; "boisei" por el antropólogo Charles Boise; "zinj", una antigua palabra para designar a África Oriental, y "anthropus", hombre. Otro cráneo fue encontrado en 1969 por Richard Leakey en Koobi Fora, cerca del lago Turkana.
Su capacidad craneal era de alrededor de 515 cm3. La cara está muy reforzada, con unos incisivos muy pequeños, pero unos enormes molares y una cresta sagital a la que debían unirse unos grandes músculos masticadores. Su foramen magnum está más adelantado que en los Australopithecus (como en el género Homo). Por lo demás, el peso, estatura y aspecto general es muy parecido a los Australopithecus.
Paranthropus boisei vivió en un medio ambiente más seco que los Australopithecus, aunque no parece que fuera un ambiente pleno de sabana. Ante el cambio climático, las especies de éste género recurrieron a la especialización de su aparato masticador para poder sobrevivir en un medio más seco. Así, con unas poderosas mandíbulas pudo tener acceso a raíces, tallos gruesos, etc. En cambio, el resto de su estructura corporal no varió prácticamente nada con respecto a sus antecesores Australopithecus.

Es el claro descendiente del Paranthropus aethiopicus. Por último, hay que destacar que el Paranthropus boisei convivió con al menos tres de las primeras especies del género Homo: Homo habilis, Homo rudolfensis y Homo ergaster.
2.2.7.3. Paranthropus robustus.
Paranthropus robustus es un hominino fósil que vivió en Sudáfrica hace entre 2 y 1,2 millones de años. Fue la primera especie descubierta del género Paranthropus, aunque en un principio se consideró perteneciente al género Australopithecus.
La denominación de robustus se debe a que los primeros hallazgos, en el sur de África, eran restos de mandíbula de gran tamaño, lo que hizo pensar que el resto del cuerpo sería enorme. Pero los descubrimientos han rechazado esta teoría, y Paranthropus robustus tenía una corpulencia similar a la de sus antepasados Australopithecus.
Tenía un gran aparato masticador, que antes se creía que era producto de una especialización alimenticia en raíces y semillas. Pero estudios recientes indican que su alimentación habría sido más variada, como diversos tipos de pastos, semillas y, posiblemente, animales.
Su rostro es achatado, con mejillas más abultadas y mandíbulas menos prominentes que el Australopithecus afarensis. Posee una pequeña cresta ósea, menor que la del Paranthropus boisei, en la parte superior del cráneo.
Tiene un marcado dimorfismo sexual, sobre todo en el desarrollo de las crestas, ausentes o muy poco marcadas en las hembras. Los machos pesaban unos 40 kilogramos y las hembras alrededor de 30 kilogramos. En cuanto a su estatura, también había grandes diferencias: el macho mediría alrededor de 1,35 metros y las hembras 1,10 metros.
La especie Paranthropus robustus sólo ha sido hallada en Sudáfrica, y su especialización parece ser menor que la de su primo el Paranthropus boisei, quizás porque no vivió en medios tan secos como aquél.
Los principales yacimientos de esta especie son los de Kromdraai, Swartkrans, Drimolen, Gondolin y Coopers. En la gruta de Swartkrans se hallaron los restos de 130 individuos; el estudio de su dentición reveló que la edad media de estos homininos era de sólo 17 años.
2.3. Los primeros Homo.
No se sabe con certeza qué especie originó los primeros miembros del género Homo; se han propuesto a Australopithecus africanus, Australopithecus afarensis y Australopithecus garhi, pero no hay un acuerdo general. También se ha sugerido que Kenyanthropus platyops pudo ser el antepasado de los primeros Homo.
Clásicamente se consideran como perteneciente al género Homo los homininos capaces de elaborar herramientas de piedra. No obstante, esta visión ha sido puesta en duda en los últimos años; por ejemplo, se ha sugerido que Australopithecus ghari, hace 2,5 millones de años, fue capaz de fabricar herramientas. Las primeras herramientas eran muy simples y se encuadran en la industria lítica conocida como Olduvayense o Modo Técnico 1. Las más antiguas proceden de la región de Afar (Etiopía) y su antigüedad se estima en unos 2,6 millones de años, pero no existen fósiles de homininos asociados a ellos.
De esta fase se han descrito dos especies, Homo habilis y Homo rudolfensis, que habitaron África Oriental entre 2,5 y 1,8 millones de años atrás, que a veces se reúnen en una sola. El volumen craneal de estas especies oscila entre 650 y 800 cm3.
2.3.1. Homo habilis.
Homo habilis es la especie más antigua del género Homo. Vivió aproximadamente desde 2,5 hasta unos 1,44 millones de años antes del presente, al comienzo del Pleistoceno. El descubrimiento de esta especie se debe a Mary y Louise Leakey, quienes encontraron los fósiles en Tanzania entre 1962 y 1964. Como se ha dicho, está generalmente aceptado que Homo habilis es la primera especie del género Homo, si bien hoy día algunos ponen en duda su pertenencia al género y ubican a Homo rudolfensis en el primer lugar.
Su nombre significa "hombre habilidoso" y hace referencia al hallazgo de instrumentos líticos confeccionados por éste. Se han realizado estudios detallados de los restos óseos de sus manos para verificar si realmente sería posible que este Homo los hubiera realizado. Los científicos concluyeron que era capaz de prensión de agarre para realizar las manipulaciones necesarias en la fabricación de utensilios de piedra. Probablemente era carnívoro oportunista. Se observa en ellos un importante incremento en el tamaño cerebral con respecto a Australopithecus, que se ha calculado entre 650 y 800 cm3.

Los restos se han hallado en Kenia, en la localidad de Koobi Fora y en Tanzania, en la conocida Garganta de Olduvai.
Las principales características son las siguientes:
- Cráneo más redondeado.
- Incisivos espadiformes.
- Molares grandes y con esmalte grueso.
- Ausencia de diastema.
- Foramen magnum ubicado más hacia el centro.
- Rostro menos prognato que los australopitecinos.
- Incisivos más grandes que los australopitecinos.
- Cara corta.
- Dedos curvos de pies y manos, lo que indica que aún utilizaban los árboles.
- La posición bípeda en las hembras provoca una reducción de la pelvis que tiene como consecuencia un adelanto de los partos (lo cual implica prematuración del neonato, un mayor tiempo dedicado a la crianza y así la necesidad de mantener vínculos sociales fuertes que potencialmente coadyuvan al desarrollo de una cultura).
- Mayor capacidad neurocraneana.
Analizando algunos de los restos óseos recuperados se puede reconocer que se trata de una especie con un aspecto mucho más humano que el encontrado en los australopitecinos. La cabeza del fémur es más grande, corta y redondeada. También la pelvis tiene un aspecto más moderno. Su altura era similar a Australopithecus africanus, de unos 1,3 metros, así como su peso, calculado en 40 kilogramos, aproximadamente.
Hallazgos realizados en el noreste africano (zona del lago Turkana) por Louise y Meave Leakey publicados en 2007 aproximan la existencia del Homo habilis hasta por lo menos 1.440.000 años antes del presente. Tal datación implica que al menos por unos 500.000 años coexistieron en los mismos territorios Homo habilis y Homo erectus. Las investigadoras opinan que inicialmente no debieron haber existido mayores conflictos entre las dos especies, sin embargo el crecimiento de la población de Homo erectus habría terminado con una lucha en pos de los recursos de la cual habrían salido exitosos los Homo erectus. Por otra parte, ese mismo hallazgo pone en duda una filiación directa entre ambas especies, aunque hay autores como Erik Trinkaus que opinan que la convivencia no descarta que Homo habilis sea ancestro directo de Homo erectus.
2.3.2. Homo rudolfensis.
Homo rudolfensis es una especie de hominino fósil, propuesta por Valerii Alexeev en 1986, cuyo espécimen tipo es el KNM-ER 1470, encontrado en Koobi Fora (orilla oriental del lago Turkana), por Bernard Ngeneo, un miembro de equipo de Richard Leakey, en 1972. Alexeev lo designó en 1986 como Pithecanthropus rudolfensis, aunque quienes lo consideran una especie distinta lo asignan al genero Homo o al género Australopithecus.
Algunos paleoantropólogos dudan de que sea una especie diferente de Homo habilis, pero no es ésta la opinión dominante en la actualidad, debido a unas marcadas diferencias morfológicas entre ambos tipos, entre las que hay que distinguir las siguientes:
- Forma de la cara, principalmente en la región supraorbital y malar, que presenta muy larga, profunda e inclinada hacia delante.
- Medidas craneales en su conjunto: Un 45% de las medidas que se compararon entre las dos especies superan el dimorfismo sexual de los gorilas.
- Volumen craneal: Alrededor de los 750 cm3, frente a los 650 cm3 del Homo habilis.
- Dentadura: Unos dientes postcaninos más amplios, con raíces y coronas más complejas y esmalte más grueso.
El Homo rudolfensis habitó en un periodo de tiempo comprendido entre 2,4 (edad del espécimen fósil UR 501) y 1,9 millones de años en el este de África, por lo que se considera contemporáneo del Homo habilis, así como de otros homininos, como el Paranthropus boisei. Su área de dispersión parece haber sido exclusivamente el este africano, por lo que muchos paleoantropólogos lo consideran un endemismo.
A partir del descubrimiento del Kenyanthropus platyops, se acepta que Homo rudolfensis podría haber evolucionado a partir de Kenyanthropus, si bien existen otras hipótesis sobre sus relaciones filogenéticas.
Finalmente, el hecho de la coexistencia de estas dos especies africanas (Homo rudolfensis y Homo habilis), ha llevado a algunos autores a sugerir un interesante debate alrededor de la coexistencia de dos especies inteligentes, pues parece estar clara la asociación de Homo habilis con las industrias líticas de Olduvai y Homo rudolfensis con las del lago Turkana, que todavía no ha llegado a sus últimas consecuencias.
El hallazgo de dos fósiles, un hueso de mandíbula de Homo habilis de 1,44 millones de años y un cráneo de Homo erectus de hace 1,55 millones de años, en el norte de Kenia, por un equipo de expertos liderado por Meave y Louise Leakey, además de demostrar que ambas especies coexistieron, ha dado nuevos argumentos para sostener la hipótesis según la cual Homo erectus desciende de Homo rudolfensis. Algunos expertos sostienen que Homo erectus y Homo rudolfensis fueron especies hermanas descendientes de alguna otra que vivió en África, de 2 a 3 millones de años antes del presente.
2.4. El poblamiento de Eurasia.
Esta es sin duda la etapa más confusa y compleja de la evolución humana. El sucesor cronológico de los citados Homo rudolfensis y Homo habilis es Homo ergaster, cuyos fósiles más antiguos datan de hace aproximadamente 1,8 millones de años, y su volumen craneal oscila entre 850 y 880 cm3.
Morfológicamente es muy similar a Homo erectus y en ocasiones se alude a él como Homo erectus africano. Se supone que fue el primero de nuestros antepasados en abandonar África; se han hallado fósiles asimilables a Homo ergaster (o tal vez a Homo habilis) en Dmanisi (Georgia), datados en 1,8 millones de años de antigüedad y que se han denominado Homo georgicus, que prueban la temprana salida de África de nuestros antepasados remotos.
Los científicos pensaban antes que los avances en la fabricación de utensilios de piedra podían haber permitido a los hombres primitivos desplazarse a Asia y Europa, tal vez ayudándoles a buscar nuevos tipos de alimentos como la carne de mamíferos grandes. Si las poblaciones humanas africanas hubieran desarrollado utensilios que les hubieran permitido capturar de forma eficaz caza mayor, hubieran tenido una fuente segura de alimentación en cualquier lugar. Según esta hipótesis, el hombre emigró a Eurasia gracias a una adaptación cultural única.
Hace unos 1,6 millones de años, los homininos habían comenzado a fabricar nuevos tipos de utensilios del tipo Achelense. Entre los utensilios achelenses comunes se encuentran grandes hachas de mano y hendedores. Aunque estas nuevas herramientas pudieron haber ayudado en la caza, los primeros utensilios achelenses conocidos en África son posteriores a la primera presencia humana conocida en Asia. Asimismo, la mayoría de los yacimientos del este de Asia de más de 200.000 años de antigüedad sólo contienen guijarros y lascas de factura sencilla (Olduvayense). Por el contrario, los utensilios achelenses eran de factura más elaborada, de mayor tamaño y más simétricos. Por tanto, los primeros colonos de Eurasia no tenían una tecnología verdaderamente Achelense y únicamente el avance en la fabricación de utensilios no puede explicar la dispersión fuera de África.
Otra posibilidad es que la primera dispersión de hombres hacia Eurasia no fue la única, sino que más bien formó parte de una migración más amplia de animales carnívoros, tales como leones y hienas. La migración humana fuera de África tuvo lugar durante la primera parte del Pleistoceno, aproximadamente hace entre 1,6 millones y 780.000 años. Muchos carnívoros africanos emigraron a Eurasia durante el comienzo del Pleistoceno y puede que el hombre también emigrara con ellos. Según esta hipótesis, el hombre fue una de las muchas especies de carnívoros que se dispersaron por Eurasia procedentes de África, y no una especie adaptada de forma singular. El hecho de depender de la carne como fuente de alimentación fundamental puede haber permitido a muchas especies carnívoras, incluido el hombre, desplazarse a través de entornos diferentes sin necesidad de tener que aprender rápidamente a distinguir entre plantas desconocidas las posiblemente venenosas.
Sin embargo, la migración humana hacia el este de Asia pudo haber tenido lugar de forma gradual y a través de latitudes inferiores y entornos similares a los de África. Suponiendo que las poblaciones del este africano se desplazasen a una velocidad de sólo 1,6 kilómetros cada 20 años, podían haber alcanzado el Sudeste Asiático en 150.000 años. A lo largo de todo este tiempo el hombre pudo haber aprendido a reconocer las plantas comestibles. Así, el hecho de comer carne puede no haber desempeñado un papel crucial en las primeras migraciones humanas hacia nuevos continentes. La comparación entre fósiles de animales, utensilios de piedra y primeros fósiles humanos procedentes de África, Asia y Europa ayudará a los científicos a establecer con mayor exactitud los factores que impulsaron y permitieron al ser humano aventurarse fuera de África por primera vez.
Esta primera migración humana condujo a la diferenciación de dos linajes descendientes de Homo ergaster: Homo erectus en Extremo Oriente (China, Java) y Homo antecessor/Homo cepranensis en Europa (España, Italia). Por su parte, los miembros de Homo ergaster que permanecieron en África inventaron un modo nuevo de tallar la piedra, más elaborado, denominado Achelense o Modo Técnico 2 (hace 1,6 ó 1,7 millones de años). Se ha especulado que los clanes poseedores de la nueva tecnología habrían ocupado los entornos más favorables desplazando a los tecnológicamente menos avanzados, que se vieron obligados a emigrar. Ciertamente sorprende el hecho que Homo antecessor y Homo erectus siguieran utilizando el primitivo Modo Técnico 1 (Olduvayense) cientos de miles de años después del descubrimiento del Achelense. Una explicación alternativa es que la migración se produjera antes de la aparición del Achelense.
Parece que el flujo genético entre las poblaciones africanas, asiáticas y europeas de esta época fue escaso o nulo. Homo erectus pobló Asia Oriental hasta hace sólo unos 50.000 años (yacimientos del río Solo en Java) y que pudo diferenciar especies independientes en condiciones de aislamiento (como Homo floresiensis en la isla de Flores, Indonesia). Por su parte, en Europa se tiene constancia de la presencia humana desde hace casi un millón de años (Homo antecessor), pero se han hallado herramientas de piedra más antiguas no asociadas a restos fósiles en diversos lugares. La posición central de Homo antecessor como antepasado común de Homo neanderthalensis y Homo sapiens ha sido descartada por los propios descubridores de los restos (Eudald Carbonell y Juan Luis Arsuaga).
Los últimos representantes de esta fase de nuestra evolución son Homo heidelbergensis en Europa, que supuestamente está en la línea evolutiva de los neandertales, y Homo rhodesiensis en África que sería el antepasado del hombre moderno.
Una visión más conservadora de esta etapa de la evolución humana reduce todas las especies mencionadas a una, Homo erectus, que es considerada como una especie politípica de amplia dispersión con numerosas subespecies y poblaciones interfértiles genéticamente interconectadas.
2.4.1. Homo ergaster.
Homo ergaster es un hominino extinto ya plenamente humano. Se estima que apareció hace unos 2 millones de años, y desapareció hace un millón de años.
Sus primeros restos fueron encontrados en 1975 en Koobi Fora (Kenia); se trata de, al menos, dos cráneos (KNM-ER 3733, tal vez femenino, y KNM-ER 3883) de hace 1,75 millones de años, cuyo cerebro tenía un tamaño estimado en unos 850 cm3. Luego, en 1984, fue descubierto en Nariokotome, cerca al lago Turkana (Kenia), el esqueleto completo de un niño de unos 11 años, de 1,60 metros de estatura y cerebro de 880 cm3, con una antigüedad de 1,6 millones de años; se conoce como el Niño de Nariokotome o Niño de Turkana.

Homo ergaster procede probablemente de Homo habilis y es básicamente la versión africana de Homo erectus, del que es antecesor. Algunos especialistas consideran que pueden haber sido una única especie, debido a su gran parecido anatómico, en cuyo caso tendría prioridad su denominación como Homo erectus, pero parece asentarse la aceptación de dos especies diferentes.
Homo ergaster tiene un cráneo menos robusto y con un torus supraorbitario menos acusado que los Homo erectus asiáticos, y se asocia, en sus comienzos, a la industria lítica Olduvayense o Modo Técnico 1, para luego pasar a la Achelense o Modo Técnico 2.
A excepción del cráneo, tanto Homo erectus como Homo ergaster tienen una constitución física bastante parecida a la del hombre actual, siendo similar en estatura, aunque en general son de complexión más robusta y fuerte, y tienden a ser algo más anchos de caderas. Las proporciones de piernas y brazos son también totalmente modernas.
Se especula que Homo ergaster pueda ser el primer hominino con capacidad para el lenguaje articulado; sin embargo la estructura de las vértebras del Niño de Turkana parecen refutar tal hipótesis. Dentro de las hipótesis muy probables, se considera que los Homo ergaster pueden haber sido los primeros homininos en establecer relaciones sociales muy complejas, precisamente facilitadas por los primordios de lenguaje oral articulado que se veía acompañado por un gran cerebro capacitado ya para elaborar rudimentarias abstracciones (como metáforas: si observaban la huella de un animal es muy probable que rápidamente pudieran imaginar a qué animal correspondía tal huella, esto posibilitado por el suficiente desarrollo de las áreas corticales prefrontales y frontales del cerebro junto con las del lenguaje). Los artefactos de la ya mencionada industria lítica correspondientes a los Homo ergaster son muy refinados. También se supone que son los primeros ancestros de la humanidad cuya esclerótica blanca resultaba ya muy notoria, haciendo contraste con las pupilas; si esto ocurría es también probable que cada individuo Homo ergaster pudiera establecer una primitiva teoría de la mente por la cual podría intuir los estados afectivos o anímicos de sus congéneres observando las miradas de los mismos. Sin embargo también se considera que no estaban capacitados para tener una imaginación que les hiciera proyectar pensamientos de futuro a largo plazo (de hecho, el promedio de vida de Homo ergaster habría de ser escaso y poquísimos individuos superarían los 20 años de edad).
2.4.2. Homo georgicus.
Homo georgicus es un especie de hominino establecida en 2002 a partir de los fósiles encontrados un año antes en Dmanisi, en el Cáucaso, República de Georgia. Se considera intermedia entre el Homo habilis y el Homo erectus y relacionada con el Homo ergaster. Los fósiles se han datado en 1,8 millones de años. El tamaño del cerebro se ha calculado entre 600 y 680 cm3. La estatura se ha estimado en 1,5 metros.
Fue encontrado primero gran parte de un esqueleto. Posteriormente han habido otros tres hallazgos, incluido un cráneo completo (pero sin dientes, sólo con el canino izquierdo) y además, se han encontrado asociados con los huesos artefactos de piedra y de percusión, que le permitían a esta especie cazar, matar animales y procesarlos, a partir de los cuales ha sido establecida la condición de cazador y no de carroñero ni de simple recolector y consumidor de alimentos blandos vegetales del Homo georgicus. El hominino de Dmanisi consumía carne, y este producto pudo haber sido la clave de la supervivencia de esta especie y de otros homininos habitantes de altas latitudes, sobre todo en invierno.
Al parecer los cinco individuos del Dmanisi formaban una familia que se vio envuelta en una erupción volcánica que los obligó a penetrar en el abrigo, y luego murieron por asfixia debido a las cenizas volcánicas. Esa es la explicación que se desprende del estudio de los estratos, y la razón de por qué en este caso excepcional se encontró un número elevado de fósiles juntos de edades tan variadas.
2.4.3. Homo erectus.
El Homo erectus es un hominino extinto que vivió entre 1,8 millones de años y al menos 300.000 años antes del presente (Pleistoceno Inferior y Medio). Emigró desde África y habitó en Europa, China y alcanzó Indonesia.

Era muy robusto y tenía una talla elevada, casi 1,80 metros de media. El volumen craneal era muy variable, entre 800 y 1.200 cm³ (la media fue aumentando a lo largo de su dilatada historia). Poseía un marcado torus supraorbitario y una fuerte mandíbula sin mentón, pero de dientes relativamente pequeños. Presentaba un mayor dimorfismo sexual que el hombre moderno. La industria lítica que producía pertenece principalmente al Achelense y probablemente conocía el uso del fuego.
Entre 1891 y 1892 el médico anatomista neerlandés Eugène Dubois creyó encontrar el «eslabón perdido» hipotetizado por Charles Darwin al descubrir algunos dientes sueltos, una calota craneal y un fémur (muy similar al del hombre moderno) en las excavaciones paleontológicas que realizaba en el río Solo cerca de Trinil, en el interior de la isla de Java (Indonesia). Dubois publicó estos hallazgos con el nombre de Pithecanthropus erectus ("hombre-mono erguido") en 1894, pero más conocido popularmente como el Hombre de Java u Hombre de Trinil. No será hasta 1940 cuando se atribuirán estos restos al género Homo (Homo erectus erectus).
En 1921 el geólogo sueco Gunnar Anderson descubrió un molar superior y un premolar inferior humanos, en el yacimiento de Zhoukoudian, China. Desde 1926 el anatomista canadiense Davidson Black dirigió un equipo de investigación que realizó exitosas excavaciones en la zona. En 1927 fue encontrado un molar inferior con el que Black propuso la especie Sinanthropus pekinensis. En 1928 se encontraron dos cráneos y Black mostró cómo el Sinanthropus debía estar emparentado con el Pithecanthropus de Java. En la década de 1930, el anatomista y antropólogo físico alemán Franz Weidenreich y otros expertos completaron el descubrimiento de la colección más famosas de fósiles de Homo erectus, cerca de Pekín, fósiles que en su conjunto recibieron el nombre popular de Hombre de Pekín. Weidenreich fue quien reinterpretó en 1940 estos restos como Homo erectus pekinensis, una subespecie de Homo erectus.
En la cueva de Zhoukoudian se encontraron restos de más de 30 individuos que habían vivido entre hace 500.000 y 250.000 años. Estos fósiles, 14 cráneos, 14 mandíbulas inferiores, 148 piezas dentales y algunos restos postcraneales, se perdieron en 1941, al comienzo de la II Guerra Mundial, pero Weidenreich había hecho unas réplicas excelentes de los descubrimientos.
En excavaciones posteriores en este yacimiento se encontraron más restos de Homo erectus. En 1959 fue encontrada una mandíbula completa y en 1966 se hallaron fragmentos de huesos occipitales y frontales que concordaban con modelos de cráneos encontrados antes. Las excavaciones se multiplicaron después de 1972.
Miles de instrumentos líticos han sido encontrados en asociación con fósiles del Hombre de Pekín: tajadores, rascadores, cuchillos, martillos, yunques y algunas puntas. También instrumentos de huesos de ciervo, gamo y búfalo, como cavadores, cuencas, cuchillos y puntas. Se ha comprobado además que el Hombre de Pekín usaba el fuego para cocinar, pues en su cueva se han encontrado cenizas y carbón vegetal asociados a huesos de animales y a semillas quemadas; sin embargo, no hay acuerdo sobre si era fuego conservado de fuentes naturales o era ya producido artificialmente.

En China se encontraron otros yacimientos importantes de fósiles de esta especie como, por ejemplo, Lantian, Yuanmou, Yunxian y Hexian. Los investigadores también han encontrado gran número de utensilios fabricados por el Homo erectus en yacimientos como Nihewan y Bose, en China, y en otros lugares de antigüedad similar (al menos entre un millón y 250.000 años de antigüedad).
Luego se descubrió en Kenia el Homo ergaster, que se puede considerar el Homo erectus africano y probablemente la especie original, y en Dmanisi, República de Georgia, en el Cáucaso, el Homo georgicus, camino hacia el Homo erectus de Extremo Oriente, pero relacionado descendiente del Homo habilis y posiblemente del Homo ergaster, con lo cual se dibujó la ruta que siguieron los homininos que dejaron África hasta dispersarse por Asia.
El conjunto de estos y otros hallazgos es clasificado actualmente dentro del género Homo y son designadas la especies de los Hombres de Java (Hombre de Trinil) y Pekín como Homo erectus, que parece haber evolucionado en África como Homo ergaster, a partir de las poblaciones anteriores de Homo habilis, para a continuación dispersarse por gran parte de Asia desde hace unos 1,7 millones de años.
Los últimos fósiles conocidos de la especie Homo erectus, procedentes del río Solo, en Java, están datados en unos 130.000 a 50.000 años antes del presente (datación que puede ser discutible). De esta forma el Homo erectus fue una especie de gran éxito: se dispersó ampliamente y gozó de larga vida.
Desde el descubrimiento del Homo erectus, los científicos se preguntan si esta especie era un antepasado directo del Homo sapiens, debido a que las investigaciones hechas no eran suficientes para demostrarlo. Las últimas poblaciones de Homo erectus (tales como las del río Solo en Java) pueden haber vivido hace solamente 50.000 años, simultáneamente con poblaciones de Homo sapiens, y se descarta que a partir de estas últimas poblaciones de Homo erectus haya evolucionado el Homo sapiens. Aunque poblaciones anteriores de Homo erectus asiáticos podrían haber dado lugar al Homo sapiens, hoy se considera más probable que el Homo sapiens hubiera evolucionado en África probablemente de poblaciones africanas de Homo erectus, y luego los primeros Homo sapiens habrían migrado desde el noreste de África hace menos de 100.000 años al Asia, donde tal vez se encontró con los últimos Homo erectus.
En cuanto a la filogenia Homo habilis/Homo erectus, aunque es considerada posible, no parece probable de un modo directo (existiría con más probabilidad un nexo de estas especies con el Homo rudolfensis); lo concreto, como ya se ha mencionado anteriormente, es que los hallazgos realizados en la zona del lago Turkana por Louise y Meave Leakey indican que los Homo habilis vivieron en África hasta hace al menos 1.440.000 años, lo que significa que ambas especies coexistieron por un lapso de unos 500.000 años.

|
| Ver vídeo complementario sobre Homo erectus y las herramientas líticas. [9:57] |
Una especie que aparentemente desciende de forma tardía del Homo erectus es el pequeño Homo floresiensis.
2.4.4. Homo floresiensis.
Homo floresiensis (Hombre de Flores) es el nombre propuesto para una posible especie recientemente descrita del género Homo, extraordinaria por el pequeño tamaño de su cuerpo y su cerebro, y por su reciente supervivencia, pues se cree que fue contemporánea con los humanos modernos en la isla indonesia de Flores. Se descubrió un esqueleto subfósil, datado hace 18.000 años, muy completo excepto por los huesos del brazo, que todavía no se habían encontrado, en yacimientos en la cueva de Ling Bua en 2003. Posteriormente, en el mismo lugar se recuperaron partes de otros seis individuos, todos diminutos, así como los huesos del brazo derecho pertenecientes al ejemplar original y herramientas de piedra igualmente pequeñas de estratos comprendidos de entre 90.000 a 13.000 años de antigüedad.
La isla de Flores ha sido descrita como «una especie de mundo perdido», donde animales arcaicos, largamente extintos en el resto del mundo, habían evolucionado a formas gigantes y enanas por especiación alopátrica. La isla tenía elefantes enanos (una especie del género Stegodon) y lagartos gigantes similares al dragón de Komodo, así como el Homo floresiensis, que puede ser considerado una especie de humano enano.
Los descubridores han apodado a los miembros de la diminuta especie «hobbits», como la raza ficticia de Tolkien que en sus libros eran aproximadamente de la misma estatura. Igualmente hace recordar a seres de la mitología de la isla, que presenta abundantes referencias a Ebu Gogo, un hombre peludo y pequeño.
Este descubrimiento ha sido objeto de una gran controversia en el mundo científico, ya que no todos los investigadores están a favor del descubrimiento de una nueva especie, y se han publicado investigaciones a favor y en contra del Homo floresiensis.
Los primeros (y por ahora únicos) especímenes fueron descubiertos por un equipo conjunto de paleoantropólogos australianos e indonesios que buscaba en la isla de Flores evidencias de la migración humana original de Homo sapiens desde Asia hacia Australia. No esperaban encontrar una nueva especie, y quedaron bastante sorprendidos al recuperar los restos de al menos siete individuos no Homo sapiens, de entre 38.000 a 13.000 años de antigüedad, de la cueva de caliza Liang Bua en Flores. Un hueso de brazo, provisionalmente asignado al Homo floresiensis, tiene unos 74.000 años de antigüedad. También están ampliamente presentes en esta cueva sofisticados utensilios de piedra de un tamaño considerado adecuado para un humano de un metro de estatura. Dichos utensilios parecen tener una antigüedad entre 95.000 y 13.000 años y están asociados con estegodontes jóvenes, presumiblemente presas del Hombre de Flores.
Los especímenes no están fosilizados, sino que tienen lo que ha sido descrito como «la consistencia del papel mojado secándose» (una vez expuestos, los huesos deben dejarse secar antes de que puedan ser extraídos).
Los investigadores esperan encontrar ADN mitocondrial en buen estado para compararlo con muestras de especímenes no fosilizados de Homo neanderthalensis y Homo sapiens similares. Sin embargo, la probabilidad de que se haya conservado ADN es baja, pues éste se degrada más rápidamente en entornos tropicales templados, donde se sabe que no se conserva más que algunas docenas de años. La contaminación del entorno cercano parece altamente probable dado el ambiente húmedo en el que fueron encontrados los especímenes.
El Homo erectus, señalado como el antepasado inmediato del Homo floresiensis, tenía aproximadamente la misma talla que otra especie descendiente, la de los humanos modernos. Sin embargo, los especialistas creen que a consecuencia del limitado aporte alimentario de la isla de Flores, el Homo erectus, llegado a este territorio hace unos 500.000 años, sufrió un fuerte enanismo isleño (una forma de adaptación evolutiva también presente en la isla en diversas especies, incluyendo el mencionado Stegodon enano, y observada también en otras islas pequeñas). Aparte de la diferencia de tamaño, los especímenes parecen por lo demás semejantes en sus características al Homo erectus, del que se sabe vivía en el sureste asiático en la misma época que los hallazgos más antiguos del Homo floresiensis. Estas semejanzas observadas forman la base del establecimiento de la relación filogenética sugerida.
Aparte de la diferencia de tamaño, esta hipótesis se basa en que los especímenes parecen por lo demás semejantes en sus características al Homo erectus. Estas semejanzas observadas forman la base para el establecimiento de la relación filogenética sugerida. Sin embargo hay que tener en consideración que no se han encontrado en la isla restos correspondientes al Homo erectus, y mucho menos formas de transición, encontrándose hasta el momento solamente pruebas materiales (herramientas de piedra), atribuibles sólo presuntamente a una posible ocupación del Homo erectus hace 840.000 años, no descartándose también que sean atribuibles a otra posible especie de hominino existente en esa época.
Así, el espécimen tipo de esta especie encontrada, un esqueleto bastante completo, y un cráneo casi completo de una hembra de 30 años de edad y cerca de un un metro de altura, no sólo presenta una drástica reducción en comparación con el Homo erectus, sino incluso una talla algo menor que la del Australopithecus, un ancestro tres millones de años más antiguo y que no se pensaba previamente que se hubiese expandido más allá de África. Esto tiende a calificar al Homo floresiensis como el miembro más «extremo» de la extensa familia humana, ya que serían ciertamente los más bajos y pequeños.
En relación a la estatura, el Homo floresiensis es también bastante diminuto comparado con el tamaño del ser humano moderno. La altura estimada de un Homo floresiensis adulto es considerablemente menor que la altura media adulta de todas las poblaciones humanas modernas físicamente más pequeñas, tales como los pigmeos africanos (< 1,5 metros), Twa, Semang (1,37 metros para las mujeres adultas) o los andamaneses (1,37 metros para las mujeres adultas). La masa es normalmente considerada más importante biofísicamente que una medida unidimensional de altura, y por dicha medida, debido a los efectos de escala, las diferencias son incluso mayores. Se ha estimado que en el espécimen tipo de Homo floresiensis era de unos 25 kilogramos.
El Homo floresiensis además tenía brazos relativamente largos, quizás para permitir a esta pequeña criatura trepar a la seguridad de los árboles cuando lo necesitaba. Estos huesos del brazo, hacen que las inevitables comparaciones con los humanos modernos acondroplásicos (sobre 1,2 metros) u otros enanos no sean válidas, pues estas personas no son proporcionalmente más pequeñas que las demás por regla general, sino que sólo tienen sus miembros más cortos.
Además de una talla corporal pequeña, el Homo floresiensis tenía un cerebro extraordinariamente pequeño. El espécimen tipo, con 380 cm3, está en el mismo rango que los chimpancés o los antiguos Australopithecus. Su cerebro está reducido considerablemente respecto al del presunto antecesor inmediato de esta especie, el Homo erectus, que con 980 cm3 tenía más del doble de volumen cerebral que su especie descendiente. Sin embargo, la relación cerebro-masa corporal que presenta el Homo floresiensis es comparable a la del Homo erectus, lo que indica que es improbable que las especies difieran en inteligencia. De hecho, los descubridores han asociado a la especie algunos comportamientos avanzados.
Estos comportamientos estarían asociados a la existencia de evidencias del uso del fuego para cocinar. La especie también ha sido relacionada con herramientas de piedra de la sofisticada tradición del Paleolítico Superior típicamente asociada con los humanos modernos, quienes con 1.310-1.475 cm3 casi cuadruplican el volumen cerebral del Homo floresiensis (con una masa corporal incrementada en un factor de 2,6). Algunas de estas herramientas fueron aparentemente usadas en la caza necesariamente cooperativa del Stegodon enano local por esta pequeña especie humana. Igualmente en otro yacimiento llamado Mata Menge, el investigador Adam Brumm y sus colaboradores han detectado que las herramientas encontradas tienen importantes similitudes con las halladas en Liang Bua; presentando las herramientas de Mata Menge una datación que arroja la asombrosa fecha de entre 840.000 y 700.000 años. Estas fechas, indican que Homo sapiens no pudo fabricar las herramientas descubiertas en Mata Menge, y con ello probablemente también las de Liang Bua, ya que la especie Homo sapiens no existía en esa época.
Además, la isla de Flores permaneció aislada durante la edad de hielo más reciente debido a un profundo estrecho, a pesar del bajo nivel marino que unió a buena parte del resto de Sundaland. Este hecho ha llevado a los descubridores del Homo floresiensis a concluir que la especie o sus antecesores sólo pudieron haber alcanzado la isla aislada por medio de algún transporte marítimo, quizás llegando en balsas de bambú hace unos 100.000 años.
Estas evidencias observadas de tecnología avanzada y cooperación a un nivel humano moderno ha impulsado a los descubridores a proponer que Homo floresiensis tendría casi con total certeza lenguaje. Estas sugerencias han resultado ser las más controvertidas de los hallazgos de los descubridores, a pesar de la probablemente alta inteligencia del Homo floresiensis.
El otro aspecto notable del hallazgo es que se cree que esta especie ha sobrevivido en la isla de Flores tan recientemente como hasta hace 12.000 años. Eso la hace el hominino más longevo, sobreviviendo largamente la desaparición de los neandertales hace unos 28.000 años. El Homo floresiensis ciertamente coexistió con los humanos modernos, quienes llegaron a la región hace entre 35.000 y 55.000 años, durante un largo período, pero se desconoce cómo pudieron haber interactuado.
El análisis de la geología local sugiere que una erupción volcánica en Flores fue la responsable de la desaparición del Homo floresiensis en la parte de la isla bajo estudio hace aproximadamente 12.000 años, junto con gran parte de la fauna local, incluido el Stegodon enano y las ratas gigantes de la isla.
Los descubridores sospechan, sin embargo, que esta especie puede haber sobrevivido mucho más tiempo en otras partes de la isla de Flores hasta llegar a ser el origen y fuente de las historias sobre los Ebu Gogo contadas entre los lugareños. Se dice que los Ebu Gogo eran cavernícolas pequeños, peludos y de lenguaje pobre, y presentarían el tamaño del Homo floresiensis. Se creía ampliamente en su existencia en la época de la llegada de los holandeses hace quinientos años, e incluso algunas historias dicen que estas extrañas criaturas han sido vistas hace tan sólo un siglo.
De forma similar, en la isla de Sumatra circulan leyendas sobre un humanoide de un metro de altura, el Orang Pendek, aunque son pocos los investigadores profesionales que las toman en serio, recibiendo más atención de la autodenominada criptozoología (disciplina cuyo enfoque suele ser pseudocientífico). Los investigadores que han trabajado en el hombre de Flores, han señalado que el Orang Pendek y otros humanoides salvajes del folclore de Asia sudoriental podrían estar relacionados de alguna forma con relatos sobre antiguos encuentros con Homo floresiensis u otros homininos enanos aún desconocidos.
El descubrimiento es ampliamente considerado como el más importante de su clase en la historia reciente, y fue toda una sorpresa para la comunidad antropológica. La nueva especie desafía muchas de las ideas de su disciplina. Desde el siglo XIX, cuando empezaron a descubrirse los primeros neandertales, no se habían descubierto otros homininos coetáneos del Homo sapiens.
El Homo floresiensis es tan diferente en forma a otros miembros del género Homo que obliga al reconocimiento de una posible nueva e inimaginable variabilidad en ese grupo, y reafirma una tendencia intelectual lejana a la idea de la evolución lineal.

|
| Ver vídeo complementario sobre Homo floresiensis [1:50] |
2.4.5. Homo antecessor.
Homo antecessor es una especie fósil perteneciente al género Homo, considerada la especie hominina más antigua de Europa con una antigüedad de más de un millón de años (Pleistoceno Inferior). Eran individuos altos, fuertes y con una cara de rasgos modernos, aunque su cerebro fuera más pequeño que el del hombre actual.
El nombre proviene de la palabra latina antecessor que significa explorador o el que va adelante, pionero, primitivo habitante. Al asignar este nombre los autores quisieron enfatizar que los fósiles de esta especie pertenecen a la primera población de homininos que llegó al continente europeo.
La definición de esta especie es fruto de los más de ochenta restos hallados desde 1994 en el nivel TD6 del yacimiento de Gran Dolina en la Sierra de Atapuerca, España. De acuerdo con sus descubridores, entre los caracteres anatómicos de estos homininos cabe destacar un conjunto de rasgos muy primitivos en el aparato dental, que llevaron a establecer una relación entre éstos y los homininos africanos del Pleistoceno Inferior. Una mandíbula muy bien conservada de una hembra Homo antecessor, de entre 15 y 16 años, recuperada del yacimiento de la Gran Dolina tiene similitudes muy claras con las del Hombre de Pekín (Homo erectus), lo que sugiere un origen asiático de Homo antecessor. Sin embargo, el patrón de desarrollo y erupción de los dientes es prácticamente idéntico al de las poblaciones modernas.
La morfología facial es similar a la de Homo sapiens, con orientación coronal y ligera inclinación hacia atrás de la placa infraorbital que determina la presencia de una fosa canina muy conspicua. El borde inferior de esta placa es horizontal y ligeramente arqueado. El arco superciliar es en doble arco y la capacidad encefálica, estimada a partir de un fragmento incompleto de hueso frontal, indica una cifra superior a los 1.000 cm3. La morfología de la mandíbula recuerda a la de ciertos homininos muy posteriores, del Pleistoceno Medio, de la especie Homo heidelbergensis, como los de la Sima de los Huesos, también en Atapuerca. El esqueleto postcraneal indica una cierta gracilidad en comparación con la mayor robustez del Homo neanderthalensis, de la segunda mitad del Pleistoceno Medio.
En la actualidad, la validez de esta denominación como especie diferente es defendida por sus descubridores y otros expertos, que consideran que Homo antecessor precede a Homo heidelbergensis y por tanto es también antepasado de Homo neanderthalensis. Sin embargo, parte de la comunidad científica la considera una simple denominación, no específica, para referirse a restos encontrados en Atapuerca, que ellos asignan a la especie Homo heidelbergensis o bien la consideran una variedad de Homo erectus/Homo ergaster.
En marzo de 2008 se han dado a conocer nuevos restos de Homo antecessor, concretamente parte de una mandíbula de un individuo de unos 20 años y 32 herramientas de sílex de tipo Olduvayense, datados en 1,2 millones de años de antigüedad, lo que hace retroceder considerablemente la presencia de homininos en Europa. Los restos fueron hallados en 2007 en la Sima del Elefante, yacimiento situado a unos 200 metros de la Gran Dolina.
 Homo antecessor
Homo antecessorPor Juan Luis Arsuaga (1 de noviembre de 2002) [Saltarse este artículo]

En Europa hay fósiles humanos datados en unos 800.000 años en el yacimiento de la Gran Dolina en la Sierra de Atapuerca, es decir al final del Pleistoceno Inferior (que se extiende desde hace 1,7 millones de años hasta hace 780.000 años). Antes de su descubrimiento en el año 1994 no había pruebas concluyentes de presencia humana en Europa hace más de medio de millón de años. Esta ausencia de datos contrastaba con el importante número de yacimientos que se conocen a partir de esa fecha, yacimientos que se encuentran en todo el continente, incluyendo las tierras del norte, como en el caso de Boxgrove en Inglaterra, donde se ha encontrado una tibia humana y un par de dientes, y de Mauer en Alemania, de donde procede una famosa mandíbula. Otros yacimientos importantes, con algo más de 400.000 años y fósiles humanos, son los de Arago (Tautavel) en Francia y Swanscombe en Inglaterra. El utillaje lítico encontrado en estos yacimientos es el clásico del Achelense. Frente a estos testimonios incuestionables de un continente poblado en su mayor parte, no había un solo fósil humano ni objeto tallado de cronología segura de época anterior, lo que parecía sugerir que Europa fue poblada hace medio millón de años por gentes venidas de fuera con una tecnología achelense. En honor de la mandíbula de Mauer, hallada cerca de Heidelberg, a estos primeros pobladores se les dio el nombre de Homo heidelbergensis, que también se ha aplicado a otros fósiles del Pleistoceno Medio de África (Bodo, Broken Hill, Ndutu, Elandsfontein) e incluso de Asia, como los ya mencionados de Dali y Jinniushan (datados hacia el Pleistoceno Medio Final) y Yunxian (que parece más antiguo, siempre dentro del Pleistoceno Medio).
El hallazgo de los fósiles de la Gran Dolina cambió el panorama. Se encontraron en un sondeo de escasas dimensiones cerca de 80 restos humanos y de 200 piedras talladas en el nivel TD6, de polaridad magnética inversa, y que se atribuye, en base a los estudios paleontológicos, al final del cron Matuyama en la escala magnetoestratigráfica (o sea, unos 800.000 años de antigüedad). Los restos humanos pertenecen a por lo menos seis individuos diferentes de todas las clases de edad y de todas las partes del esqueleto, lo que hace pensar que quedan muchos más fósiles por descubrir. Los huesos presentan trazas de haber sido descarnados, fracturados y consumidos sin ceremonia alguna por otros humanos en un caso de canibalismo puramente alimenticio. Ésta es la más antigua prueba de tal comportamiento humano y una de las poquísimas que se conocen en la prehistoria antigua (hay un cráneo etíope de hace medio millón de años, el de Bodo, con trazas de descarnamiento, y un yacimiento mucho más reciente presenta neandertales consumidos).
A partir de la muestra de fósiles disponibles en la Gran Dolina, que se espera ampliar en unos pocos años (cuando la excavación en extensión alcance el nivel de los fósiles humanos) se ha creado la especie Homo antecessor, propuesta como el último antepasado común de neandertales y humanos modernos. Otro fósil humano que podría ser de la misma edad y de la misma especie es la calvaria incompleta hallada en Ceprano (Italia), aunque carece de contexto geológico claro y no lo tiene arqueológico ni paleontológico. Entre los yacimientos arqueológicos de hace mucho más de medio millón de años se puede mencionar el de Monte Poggiolo, en Italia. En Fuentenueva 3 y Barranco del León, en Orce (Granada), se han encontrado piedras talladas que podrían tener 1,2 millones de años, y en la cueva de la Sima del Elefante, en la Sierra de Atapuerca, hay otras algo más modernas. Sin embargo, en todos estos yacimientos es preciso confirmar los indicios, muy alentadores, con más pruebas.
A pesar de los recientes hallazgos sigue siendo real la escasez de yacimientos para hace más de medio millón de años, en contraposición con lo que se encuentra después. Además, los datos más antiguos proceden del sur de Europa, lo que se podría tomar como indicio de una población muy poco densa y exclusivamente mediterránea. Un yacimiento tan rico en especies como el de Untermassfeld, en Alemania, datado en un millón de años, no muestra presencia humana en la región.
Por otro lado, la industria de la Gran Dolina se asigna al Modo I. Al igual que en el caso del Extremo Oriente surge la pregunta de por qué los pobladores de Eurasia en esa época no tenían una tecnología, el Achelense o Modo II, desarrollada cientos de miles de años antes en África. Una respuesta posible, pero no la única, es la de que salieron de África hace más de 1,6 millones de años. Esa salida podría ser la representada en Dmanisi, pero no hay cronologías seguras tan antiguas en Europa, ni todo el mundo las admite para China y Java.
Por otro lado, la mayor abundancia de yacimientos achelenses, y por toda Europa, hace más de medio millón de años parece indicar un sustancial aumento de la población humana, que podría entenderse como resultado de una expansión demográfica y la importación de una nueva tecnología, sin movimiento de gentes, o como una segunda colonización de Europa, que reemplazaría o se fusionaría con la anterior; y hasta es posible que se encontrasen un continente vacío cuando llegaran los achelenses hace medio millón de años.2.4.6. Homo cepranensis.
El Hombre de Ceprano es un ejemplar fósil del cráneo de un hominino, descubierto por Italo Bidittu en 1994, en la localidad de Ceprano, provincia de Frosinosne, Italia. Tras la reconstrucción de los fragmentos encontrados, realizada por el geólogo Aldo Segre y la paleontóloga Eugenia Segre Naldini, se calcula que la capacidad craneana de este espécimen podría ser de hasta 1.200 cm3, lo que significa un cerebro claramente mayor que el del Homo ergaster y el Homo erectus. Se ha estimado que este fósil data de hace por lo menos 800.000 años. Las diferencias encontradas con las especies conocidas de Homo han llevado a la definición formal de este hominino como especie diferente: Homo cepranensis.
2.4.7. Homo heidelbergensis.
El Homo heidelbergensis es una especie extinta del género Homo, que surgió hace más de 500.000 años y perduró al menos hasta hace 250.000 años (Pleistoceno Medio). Eran individuos altos (1,75 metros) y muy fuertes (llegarían a los 100 kilogramos), de grandes cráneos (1.350 cm3) muy aplanados con relación a los del hombre actual, con mandíbulas salientes y gran abertura nasal. Se le dio ese nombre porque los primeros fósiles fueron descubiertos cerca de Heidelberg, Alemania.
Es un antepasado directo del Hombre de Neandertal en Europa. Aún cuando es muy similar a los Homo sapiens arcaicos encontrados en África (por ejemplo: Homo rhodesiensis y Homo sapiens idaltu), se sabe hoy que Homo heidelbergensis no fue antepasado directo de los humanos modernos. Entre Homo antecessor y Homo neanderthalensis existió esta especie (Homo heidelbergensis). Presenta en general caracteres intermedios entre Homo erectus/Homo ergaster y Homo sapiens.

Presenta un torus occipital hendido (en el Homo erectus tal torus, o cresta, es continuo) y una gran capacidad neurocraneal. Si en los Homo erectus la dieta probablemente fue basada en el cleptoparasitismo, el carroñeo (que aportaban proteínas y grasas de buena calidad) y un régimen de alimentos vegetales, en el Homo heidelbergensis se hace patente un predominio de la dieta carnívora con evidencias de caza. La presión evolutiva para que entre los heidelbergensis se desarrollara la caza habría estado en las condiciones ecológicas del territorio que colonizaron: Europa era fría y durante seis meses existían muchos menos recursos alimenticios obtenibles de vegetales que en África, por lo que el carroñeo y el cleptoparasitismo (robar presas a los animales predadores) no aportaba suficientes nutrientes. Esto habría inducido (por selección) a la aparición de conductas sociales dedicadas a la caza: bandas de heidelbergensis se organizaban para perseguir a otros animales y ultimarlos en trampas naturales (precipicios, pantanos) o arrojándoles grandes hachas líticas bifaces e, incluso, rústicos venablos de madera aguzada.

El más antiguo de los fósiles de la especie es una mandíbula inferior encontrada por el trabajador de una mina en Mauer, cerca de Heidelberg. Posteriormente, en una cueva llamada Caune de l'Arago, en Francia, se encontraron los restos fragmentarios de una docena de individuos. Uno de los más completos es la cara y parte de la caja craneana de un individuo conocido como Hombre de Tautavel, que tiene gran parecido al cráneo del Hombre de Petralona, encontrado en una cueva en Grecia. Otros sitios donde se han hallado fósiles de esta especie son Steinheim (Alemania), Swanscombe (Inglaterra) y la Sima de los Huesos en la Sierra de Atapuerca (España), donde se encontraron 5.000 fósiles pertenecientes a un mínimo de 32 individuos, que datan de hace 400.000 años, considerados antepasados de los neandertales, restos éstos que están muy bien conservados; entre ellos destacan el cráneo número 5 (llamado popularmente Miguelón, en la imagen) que está totalmente completo, y del cual recientemente se realizaron estudios que dan cuenta de una lateralidad en el cerebro (era diestro), y una pelvis muy bien conservada de un individuo conocido popularmente como Elvis. En China se han encontrado fósiles que concuerdan con este grupo, en el sitio de Dali: un cráneo de hace 280.000 años, y un esqueleto en Jinniushan.
Hay que notar que en Terra Amata (Francia), se encontraron restos de refugios construidos durante la época de Homo heidelbergensis. Estos refugios tenían una forma ovalada, de unos ocho metros de largo y siete de ancho. Dentro del refugio se encontraron restos de ceniza.
Se han encontrado fósiles relacionados con esta especie en África en los sitios de Bodo, al noreste de Etiopía, con un ejemplar más robusto pero similar que data de hace 600.000 años, y en Kabwe, Zambia. También en lago Ndutu, Tanzania. Los huesos postcraneales concuerdan con los de Arago, e indican que ésta era una forma robusta, pero moderna. Los científicos están divididos en su interpretación sobre estos fósiles africanos. Algunos los sitúan en la especie Homo heidelbergensis. Otros piensan que los fósiles europeos y africanos pertenecen a dos especies diferentes, y sólo las poblaciones africanas, clasificadas como Homo rhodesiensis u Homo sapiens arcaico, evolucionaron hasta el Homo sapiens. Tampoco hay unanimidad sobre la interpretación del Hombre de Petralona (Grecia), que podría ser un Homo heidelbergensis tardío o más bien un Homo rhodesiensis u otro Homo sapiens arcaico que llegó a Europa desde África, por las costas del Mediterráneo.
Hace aproximadamente medio millón de años se empezaron a gestar los rasgos morfológicos que en unos cientos de miles de años dieron origen al neandertal. Los pre-neandertales u Homo heidelbergensis mostraban una creciente corpulencia física, expresada en el aumento de la talla, 1,75 a 1,80 metros y cerca de 100 kilogramos de peso (media masculina). Las mandíbulas también poseían una gran fuerza y robustez a causa del estrés al cual eran sometidas.
El conjunto de estos fósiles revelan un hominino con un cráneo más "inflado" que el de ningún Homo erectus/Homo ergaster, aunque todavía posee arcos supraciliares. La parte trasera del cráneo es más redondeada que en Homo erectus/Homo ergaster, y las mejillas son infladas, como en los neandertales, aunque la cara es más plana. Su capacidad neurocraneal no dista mucho de la del hombre moderno, rondaba los 1.350 cm3. Otro parecido con el Homo sapiens actual era su aparato fonador. Esto dio a pensar que el lenguaje, entendido de una manera quizás diferente a la actual, ya estaba presente en estos grupos.
Los utensilios asociados con los fósiles consisten básicamente en "piedras de cortar" y algunas herramientas de lascas como puntas y raspadores de madera, hueso y asta, siendo los iniciadores de esta técnica. Dentro de su tecnología encontramos el bifaz, también llamado hacha de mano; es de grandes dimensiones y está tallado por ambas caras. Su utilidad es muy diversa. Según análisis de otras piedras muestran su empleo para curtir pieles o trabajar la madera. Hace 400.000 años ya usaban rudimentarias lanzas de madera. También es muy probable que en esta época el fuego fuese utilizado y paulatinamente generalizado, ya que hay evidencias de fogatas.
Dentro de esta especie también podemos empezar a hablar sobre pequeños grupos con una cohesión social, ya que según estudios paleontológicos de diferentes restos se han hallado, por ejemplo, mandíbulas totalmente edentadas; esto indica que el anciano, tras perder su dentadura, siguió viviendo ayudado por sus congéneres. Esto se traduce en una conducta diferencial a la hora del tratamiento de sus cadáveres (Sima de los Huesos). El estudio de las dentaduras de los heidelbergensis también revela otro aspecto evolutivo de sumo interés: los molares y la dentadura definitiva recién les aparecían entre los 5 y 6 años tras ser paridos (en un chimpancé actual esto ocurre hacia los 3 años). Si se considera que un individuo no puede alimentarse por cuenta propia hasta tener bien desarrollada su dentición, esto induce a pensar que los heidelbergensis estaban caracterizados por una prematuración que implicaba un prolongado cuidado de las crías, con el posible establecimiento de fuertes relaciones afectivas entre los individuos. La existencia de estas fuertes relaciones afectivas habría cohesionado a las sociedades heidelbergensis de un modo que les capacitaba para, por ejemplo, desarrollar eficaces estrategias de caza y de supervivencia en general.
La Sima de los Huesos.
El yacimiento de la Sima de los Huesos se localiza en el interior del sistema de cavidades del karst de la Sierra de Atapuerca, de la provincia de Burgos. En esta sima, situada a unos 40 metros de profundidad y de poco más de 20 metros cuadrados, se han recuperado hasta la fecha más de 6.000 restos fósiles de Homo heidelbergensis.
En la Sima de los Huesos se han encontrado fósiles humanos de al menos 32 individuos. Un hallazgo de estas características es anormal ya que no se suelen encontrar grupos de humanos juntos, a no ser que esos cuerpos hubiesen sido transportados a la Sima por algún motivo en especial. Este hecho ha permitido estudiar un grupo poblacional completo del Pleistoceno Medio.

Gracias al estudio de los restos podemos saber bastante sobre ellos. Medían 1,75 metros los varones y 1,70 metros las hembras. Conocemos algunas de las dolencias que padecían como el fuerte desgaste dental propiciado seguramente por la ingesta de frutos o vegetales sin cocinar, pero por el contrario no se ha visto ni una sola caries. Estos hombres eran diestros y usaban palillos de dientes, que dejaron surcos entre sus piezas dentales. El individuo del cráneo 5 murió seguramente de septicemia, ya que se le ha detectado una infección bucal que al no ser paliada se generalizó. Agamenón o el cráneo 4 presenta los canales auditivos tapados, por lo que se puede inferir su sordera antigua. Se puede afirmar que en general tienen buena salud, y a excepción de los impactos que presentan los cráneos, por otra pate numerosos, no hay señales de grandes traumatismos óseos.
Los dientes hablan también de épocas de escasez de alimentos o enfermedad: casi un tercio de los individuos de la Sima pasaron por momentos de estrés por malnutrición o enfermedad. En muchos casos esto ocurría hacia los tres o cuatro años, que corresponde con el destete.

Se ha podido determinar el sexo en 18 individuos, la mitad mujeres, y también su edad. No hay menores de 3 años, pero a la Sima llegaron individuos menores de 13 años, adolescentes de entre 13 y 17 años (la edad mejor representada), y sólo tres que superan los 30 años de edad. De su modo de vida se sabe que eran cazadores-recolectores, pero no por este yacimiento, ya que en él sólo aparecen asociados restos de oso, león, lince, gato montés y comadreja. La explicación sobre la gran acumulación de cadáveres en este sitio ha sido muy debatida. Claramente la Sima no sirvió como lugar de habitación, ya que no se han encontrado restos de instrumentos ni de herbívoros que, como en los demás yacimientos, así lo indique. Se han barajado varias hipótesis, y en 1998 se confirmó una de ellas gracias al hallazgo de un hacha de mano denominada Excalibur, la única herramienta hallada en este yacimiento. Por ello el equipo de investigación de Atapuerca apoya la hipótesis de que la Sima de los Huesos es una acumulación intencional de cadáveres. Esto significa que posiblemente la Sima sea el testimonio más antiguo de un comportamiento simbólico, quizás funerario, de la historia de la humanidad.
Entre los hallazgos fósiles más importantes de este yacimiento se encuentran el ya mencionado cráneo 5, que es el más completo y mejor conservado del registro fósil mundial, una pelvis completa (en la imagen) y el esqueleto completo de un pie de Homo heidelbergensis.

|
| Ver vídeo complementario sobre el yacimiento de la Sima de los Huesos [3:12] |
2.4.8. Homo rhodesiensis.
Homo rhodesiensis es una especie de hominino fósil del género Homo, hallado por primera vez en 1921 en la localidad llamada por los ingleses Broken Hill, actualmente Kabwe, en Zambia (antigua Rhodesia del Norte, por lo que se denominó Hombre de Rhodesia). Se considera que vivió solamente en África, desde hace 600.000 hasta 160.000 años antes del presente, durante el Pleistoceno Medio.
La capacidad neurocraneal del Homo rhodesiensis era relativamente alta, entre 1.280 y 1.325 cm³. Un cráneo que podría ser el más antiguo de esta especie, hallado en Bodo, Etiopía, datado en 630.000 años antes del presente, tenía una capacidad endocraneana de 1.250 cm3. Otros fósiles con características morfológicamente similares han sido encontrados en Sudáfrica (Hombre de Saldaña), Tanzania, Marruecos y Argelia.
Morfológicamente conserva ciertos rasgos antiguos comunes al Homo erectus, Homo ergaster y Homo antecessor, tiene otros que han permitido relacionarlo con el Homo heidelbergensis y Homo neanderthalensis, pero además presenta rasgos que apuntan hacia el Homo sapiens. Por tales características intermedias ha sido comparado con el Hombre de Petralona.
2.5. Las etapas finales.
La fase final de la evolución de nuestra especie está presidida por dos especies humanas, ambas inteligentes, que durante un largo periodo convivieron y compitieron por los mismos recursos. Se trata del Hombre de Neandertal (Homo neanderthalensis) y nosotros mismos, Homo sapiens. Son en realidad dos historias paralelas que se cruzan en un momento determinado.
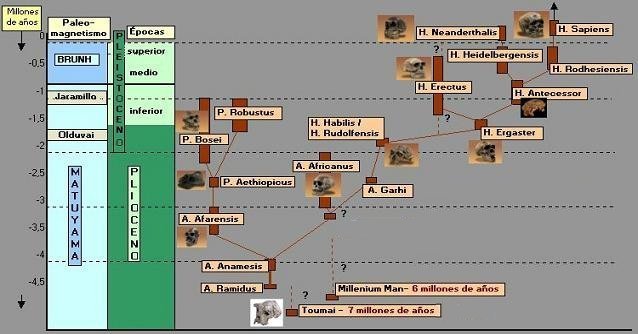
El Hombre de Neandertal surgió y evolucionó en Europa y Oriente Medio hace unos 230.000 años, presentando claras adaptaciones al clima frío de la época (complexión baja y fuerte, nariz ancha).
Los fósiles más antiguos de Homo sapiens datan de hace unos 200.000 años (Etiopía). Hace unos 90.000 años llegó a Oriente Próximo donde se encontró con el Hombre de Neandertal que huía hacia el sur de la glaciación que se abatía sobre Europa. Homo sapiens siguió su expansión y hace unos 45.000 años llegó a Europa Occidental (Francia). Paralelamente, el Hombre de Neandertal se fue retirando, empujado por Homo sapiens, a la periferia de su área de distribución (Península Ibérica, mesetas altas de Croacia), donde desapareció hace unos 28.000 años.
Aunque Homo neanderthalensis ha sido considerado con frecuencia como subespecie de Homo sapiens (Homo sapiens neanderthalensis), análisis del ADN mitocondrial de fósiles neandertales sugieren que la diferencia existente es suficiente para considerarlos como dos especies diferentes, separadas desde hace al menos 400.000 años y probablemente más.
Se tiene la casi plena certeza de que el Hombre de Neandertal no es ancestro del ser humano actual, sino una línea evolutiva paralela derivada también del Homo erectus/Homo ergaster a través del eslabón conocido como Homo heidelbergensis. El neandertal coexistió con el Homo sapiens y quizá terminó extinguido por la competencia con nuestra especie. Si existió algún mestizaje entre ambas especies, el aporte a la especie humana actual ha sido, en lo genético, inferior al 5% (aunque existe actualmente casi total escepticismo de que ambas especies hayan sido interfértiles). En cuanto al llamado Hombre de Cro-Magnon corresponde a las poblaciones de Europa Occidental de la actual especie Homo sapiens.
2.5.1. Homo neanderthalensis.
El Hombre de Neandertal, u Homo neanderthalensis, es una especie del género Homo que habitó Europa y partes de Asia Occidental desde hace 230.000 hasta 28.000 años atrás, durante el Pleistoceno Medio y Superior y culturalmente integrada en el Paleolítico Medio.
Sus características definitorias, a partir de los huesos fósiles descubiertos hasta ahora (unos 400 individuos), son: esqueleto robusto, extremidades cortas, tórax en barril, arcos supraorbitarios resaltados, frente baja e inclinada, faz prominente, mandíbulas sin mentón y gran capacidad craneal (1.500 cm3). Vivían en grupos organizados, formados por alrededor de unos treinta miembros.
Los neandertales fueron una especie bien adaptada al frío extremo. En ellos se notan cráneos alargados y amplios, complexión corta y robusta y nariz grande, rasgos que denotan adaptación a climas fríos, como se puede observar actualmente en las poblaciones del Ártico. Sus cerebros eran igual o más grandes que los del hombre moderno. Un neandertal promedio tendría una altura de 1,65 metros, de contextura pesada, y musculatura robusta.
El estilo de herramientas líticas utilizadas en el Paleolítico Medio por los neandertales es el Modo Técnico 3 o cultura Musteriense, así llamada por haber sido encontrada por primera vez en el yacimiento arqueológico Le Moustier. La cultura Musteriense está caracterizada por la utilización de la técnica de talla Levallois. Estas herramientas fueron producidas usando martillos de percusión blandos, como huesos o madera. En los últimos tiempos de los neandertales se nota en el registro arqueológico el estilo Châtelperroniense, considerado como más avanzado que el Musteriense.
2.5.1.1. Descubrimiento.

El comienzo de la historia del Hombre de Neandertal es también el inicio de la paleoantropología. En agosto de 1856, tres años antes de que Darwin publicara El origen de las especies, fue descubierto el espécimen que luego sería conocido como Neandertal 1. El lugar fue la cueva Feldhofer, en una zona encañonada del valle del río Düssel, cerca de Düsseldorf, Alemania, que se llama valle de Neander (en alemán Neanderthal) por el compositor y teólogo Joachim Neander.
En realidad, los primeros fósiles fueron encontrados en Engis (Bélgica), en 1829. Le siguen los de Gibraltar, en la cantera de Forbe en 1848, pero no se reconoció el significado de estos dos descubrimientos hasta bastante después de que se diera a conocer Neandertal 1.
El descubrimiento de 1856 fue realizado por Franz Meyer y para explicar dicho hallazgo inventó una teoría curiosa. Según Franz el esqueleto pertenecía a un cosaco ruso que perseguía a Napoleón a través de Europa. Explicaba que el cosaco sufría raquitismo, lo que explicaría la forma arqueada de sus piernas, y que el dolor del raquitismo le hacía arquear tanto las cejas que le produjeron unos fuertes arcos supraciliares.
En 1863 el geólogo inglés W. King, impresionado por la remarcable ausencia en el cráneo de Neanderthal de los contornos y proporciones que caracterizan a nuestra propia especie, creó, a partir del espécimen de Neander, una nueva especie: Homo neanderthalensis. Con la denominación propuesta por King se reconoce por primera vez formalmente la existencia en la historia biológica de la Tierra de una especie de hominino distinta de Homo sapiens.
2.5.1.2. Clasificación.
Su lugar en la clasificación científica ha sido fuertemente debatido, en ocasiones clasificándose como una subespecie de Homo sapiens (Homo sapiens neanderthalensis), pero el consenso actual lo ubica como una especie aparte (Homo neanderthalensis). La genética ha demostrado que el Hombre de Neandertal evolucionó paralelamente al Homo sapiens desde antepasados comunes lejanos. Los científicos han tenido éxito en extraer el ADN de varios fósiles de neandertales. Después de un cuidadoso análisis, particularmente del ADN mitocondrial y además de una cantidad de ADN nuclear, parece que el ADN de los neandertales es muy diferente al del Homo sapiens. Al medir el grado de diferencia filogenética entre el ADN de los neandertales y el de los humanos modernos, los investigadores sugieren que estas dos especies surgieron de linajes separados hace entre 400.000 y hasta un millón de años atrás.
Mientras el Homo sapiens evolucionó en África, desde donde migró hace entre 100.000 y 60.000 años hacia el resto del planeta, el Hombre de Neandertal se cree que desciende del Homo heidelbergensis. Estudios recientes indican que su origen podría estar en la Península Ibérica.
2.5.1.3. Existencia en el tiempo.
Las diferencias con sus antecesores no son sólo morfológicas, sino también de comportamiento: por ejemplo, practicaban enterramientos de cuerpos en compañía de alimentos, armas o utensilios. Se discute si utilizaban ciertos adornos, así como trozos de dióxido de manganeso y de ocre rojo afilados como lápices, o bien reducidos a polvo, para dibujar en las paredes.
No se han hallado restos de neandertales del periodo de la glaciación de Riss (hace 200.000-125.000), durante el cual debió producirse una lenta evolución, pero después de esta época fría, en el interglaciar Riss-Würm (hace 125.000-100.000) ya se encuentran varios restos de protoneandertales en diversos puntos de Europa, entre ellos los de Gibraltar.
En la Península Ibérica hay pruebas de su existencia desde los primeros estadios (hace unos 600.000 años) hasta hace aproximadamente 28.000 años, como indican estudios recientes.

Surgieron hace por lo menos 150.000 años en el Paleolítico Inferior y desaparecen del registro fósil hace unos 28.000 años, después de haber creado y desplegado la importante y extendida cultura Musteriense, que se considera como la expresión del Paleolítico Medio, y también el Châtelperroniense, que actualmente se cree que habría sido autóctono.
2.5.1.4. Canibalismo.
El canibalismo es algo probado en diferentes yacimientos neandertales, como Moula-Guercy o Vindija. Los restos óseos presentan marcas de corte realizadas con herramientas de piedra, y están muy bien conservados por haber sido eliminada de sus huesos toda la carne. No era un canibalismo antropofágico sino ritual, según se cree, ya que al parecer la carne se quitaba de los huesos para ser enterrados, y no para ser comida. Esto se sabe por comparación etnológica. Bajo un microscopio se nota la diferencia de los cortes realizados en animales de caza para ser comidos, y en los que se practican en huesos de los miembros fallecidos del grupo.
Esta sería una práctica que se habría realizado a lo largo de largos períodos y en diferentes regiones. Los yacimientos mencionados están el primero en Francia y el segundo en Croacia, datados en 100.000 y 30.000 años, respectivamente. También se han encontrado evidencias de canibalismo ritual en El Sidrón y Atapuerca (España), en Combe Grenal (Francia), en Krapina (Croacia) y en la cueva de Guattari (Italia), aunque en el caso de la cueva de El Sidrón se cree que podría ser un canibalismo con fines alimenticios, debido a hambrunas. Al parecer, en los dientes de esos individuos se pueden ver períodos de hambruna, y los huesos están triturados como para quitarles el tuétano.
2.5.1.5. Enterramientos.
Muchos de los esqueletos neandertales excavados en cuevas han sido históricamente considerados como el resultado de prácticas funerarias. Destacan los yacimientos de La Chapelle-aux-Saints y de La Ferrassie, éste último con ocho enterramientos (dos adultos, un niño de 10 años, dos niños de entre 2 y 3 años y tres fetos). En la cueva de Kebara se halló la mitad superior de un esqueleto neandertal, en el que tras la inhumación fue retirado el cráneo, porque se ha hallado su mandíbula y un diente de la dentición superior. Esto parece demostrar que poseían un alto nivel de sensibilidad y, de alguna forma, dedicaban un trato especial a sus muertos.
Existen mayores dudas sobre la existencia de un ritual acompañante, es decir, si el enterramiento tenía algún tipo de significado simbólico. Las pruebas de ritualidad en los enterramientos neandertales son muy dudosas:
- En el sedimento alrededor de uno de los esqueletos de Shanidar (Irak) se identificaron granos de polen procedentes de flores que, supuestamente, habían sido depositadas sobre el cuerpo.
- Sobre el esqueleto de un niño de dos años de Dederiyeh (Siria) se halló un útil triangular de sílex a la altura del corazón y, junto a la cabeza, una losa de caliza.
- En muchos otros casos, como en La Ferrassie, Amud o Teshik-Tash, los cuerpos aparecen asociados con restos faunísticos que han sido interpretados como ofrendas.
Sin embargo, algunos autores cuestionan estas pruebas de ritual y atribuyen una asociación casual a los objetos que acompañan al esqueleto. Aunque tiene la apariencia de ser intencionada, la mezcla de huesos humanos con restos de animales o útiles es siempre posible en un yacimiento donde hay abundancia de animales e instrumentos de piedra. Además, algunas de las excavaciones son antiguas y los métodos de excavación no eran los idóneos.
Pero, aunque el significado que los neandertales daban a los enterramientos y el simbolismo que les acompañaba se nos escapa, sí que podemos dar por seguro que los neandertales tenían un tratamiento especial con sus muertos, y que los enterraban intencionadamente.
El anciano de La Chapelle-aux-Saints.
El anciano de La Chapelle-aux-Saints fueron los restos que produjeron un estereotipo que tardaría años en desaparecer. Debido a la interpretación equivocada de este fósil por Marcellin Boule, el Hombre de Neandertal sería considerado un hombre de las cavernas salvaje y brutal que andaba arrastrando los pies y ni siquiera caminaba erguido. Sin embargo, a su debido tiempo, los antropólogos llegaron a comprender que lo que causaba realmente este impedimento al anciano era una severa artritis y otras deformidades. Entre las dolencias del anciano había una cadera izquierda deformada, un dedo del pie aplastado, una severa artritis en las vértebras cervicales, una costilla rota y una rodilla lesionada. Había perdido todos sus dientes, lo que había causado la reabsorción de sus soportes dentales.
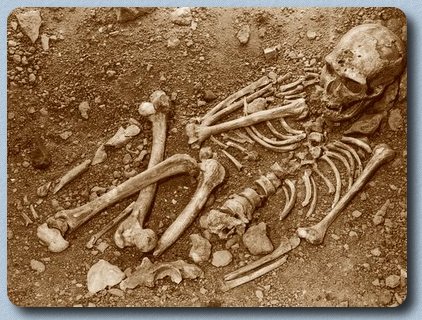
El anciano, muy parecido al neandertal de Shaindar I, probó que los neandertales cuidaban a sus ancianos, enfermos y heridos. Es evidente que el especimen de La Chapelle-aux-Saints no podía cazar ni masticar bien, ni siquiera caminar adecuadamente. Tuvo que tener algún tipo de ayuda de su grupo de parentesco para poder sobrevivir durante tanto tiempo como lo hizo.
2.5.1.6. Lenguaje.
Es polémica la cuestión de qué forma de comunicación manejaban los neandertales: si un lenguaje relativamente similar al moderno (con estructura compositiva y reglas gramaticales, de modo que un número limitado de palabras se combina para crear un número ilimitado de frases posibles) o algunas formas menos desarrolladas y, en cierto sentido, más próximas al sistema de comunicación de los simios.
Entre los autores que consideran que los neandertales no usaban un lenguaje como tal está el arqueólogo Steven Mithen, que defiende tenían un sistema de comunicación Hmmmm (esto es, holístico, manipulador, multimodal, musical y mimético). Lieberman realizó un modelo coincidente con la opinión de Mithen: la situación del cuello adelantado y la disposición de la laringe parecerían haber dificultado un lenguaje articulado. Sin embargo otros estudios suponen que el hioides estaba lo suficientemente desarrollado y posicionado como para la emisión de fonemas discretos con capacidad simbólica, aunque de un modo mucho más tosco que en el Homo sapiens.
En El Sidrón (yacimiento localizado en Asturias, de 43.000 años de antigüedad), se han podido tomar muestras que permiten reconstruir dos importantes genes neandertales: el FOXP2, relacionado con la posibilidad del habla, y del MCR1, de la pigmentación, que indicaría color del pelo rubio y pelirrojo.
2.5.1.7. Arte.
El arte de los neandertales (Musteriense) aún presenta controversias: Leroi Gouran, entre otros, observó que podían, y de hecho solían hacerlo, rendir homenajes a sus difuntos (elaborando sencillas tumbas). Bastante tardíamente, cuando ya podrían haber entrado en contacto con los Homo sapiens, los neandertales parecen haber estado dotados de la suficiente habilidad como para copiar rudimentariamente el arte de los Homo sapiens primitivos: en yacimientos correspondientes a neandertales se han hallado algunos pocos objetos de cuerno pulido que parecen haber tenido un valor estético e incluso una muy tosca máscara confeccionada con una basta placa de piedra a la cual se le practicaron dos oquedades a modo de ojos.
2.5.1.8. Extinción.
Las causas de la extinción del Hombre de Neandertal son todavía motivo de debate: ¿Los sapiens compitieron intensamente con ellos por recursos?, ¿los mataron y exterminaron en combate?, ¿les contagiaron enfermedades para las cuales carecían de defensa? o ¿no soportaron los neandertales determinados cambios climáticos o ambientales? También se especula con la posibilidad de la hibridación, o sea, que haya existido un cruce entre las dos especies y los Homo sapiens los hayan absorbido.
La hipótesis de mixogénesis Homo sapiens/Homo neanderthalensis resulta, por los mapeos de secuencias de ADN, prácticamente descartada; la extinción masiva de Homo neanderthalensis por la rigurosidad de la última gran glaciación hasta la fecha también parece descartada ya que los neandertales habrían estado muy bien adaptados al clima glacial, de modo que lo más probable (reforzada tal probabilidad por la rápida desaparición de los neandertales tras la irrupción de los Homo sapiens en Europa) es que el principal motivo para la extinción fuera la competencia con los Homo sapiens. Los últimos reductos de neandertales se encontraron en el sur de territorios actualmente españoles (Andalucía).
El niño de Lapedo.
En 1999 João Zilhão, Erik Trinkaus y otros publicaron el hallazgo en Lagar Velho (Portugal) del conocido como Niño de Lapedo, al que describían como un híbrido de Homo sapiens y neandertal porque tenía características comunes a ambos. En el mismo número de la revista, Ian Tatersall y Jeffrey H. Schwartz contestaron negando tal carácter de híbrido.

Los restos del Niño de Lapedo tienen dos problemas intrínsecos y otros dos extrínsecos. Los problemas intrínsecos son:
- Es el esqueleto de un niño, lo que siempre resulta complejo por las diferencias anatómicas entre niños y adultos (más aún cuando la base de la hipótesis consiste precisamente en las proporciones de los huesos).
- Por desgracia, el cráneo quedó destruido por la excavadora que cavaba la zanja en la que se encontró. Así pues, lo que más hubiera contribuido a establecer la supuesta hibridación entre neandertales y sapiens había desaparecido.
Los problemas extrínsecos son:
- La datación por carbono-14 de restos orgánicos relacionados con el esqueleto arrojó un resultado de 24.500 años (los restos de Homo neanderthalensis más recientes, los del Boquete de Zafarralla, están datados en 28.000 años, por lo que ni siquiera hay pruebas de que en ese momento existieran neandertales).
- El problema genético: En 1997 Krings et al. lograron obtener ADN de unos restos óseos neandertales. En 2000 Ovchinnikov et al. hicieron lo propio con otro ejemplar. El resultado de ambos es que dentro de los neandertales había una gran diversidad genética, pero si bien ambas muestran eran diferentes entre sí, lo eran aún en mayor medida (y más de lo esperado) con los Homo sapiens. Somos especies diferentes que divergieron más tempranamente de lo que se creía. Como especies diferentes, no había hibridación posible.
Los mestizosPor Juan Luis Arsuaga (1 de julio de 2002) [Saltarse este artículo]
El estatus taxonómico de los neandertales también había ido cambiando en esos mismos años. En la época anterior de simplificación taxonómica de la filogenia humana, los neandertales no eran reconocidos como una especie diferente de la nuestra, el Homo sapiens, sino tan sólo como una subespecie: Homo sapiens neanderthalensis. Pero soplaron nuevos vientos en el mundo de la paleoantropología, y el número de especies de homínidos se multiplicó. Se abandonaban así los esquemas lineales de la evolución humana, y se optaba por la geometría ramificada que es común en el resto de grupos biológicos. A este nuevo enfoque contribuyeron también el auge en paleontología humana de la cladística, una nueva metodología sistemática, y la irrupción en este campo de la escuela evolutiva llamada "teoría del equilibrio puntuado", que había nacido en la década de los setenta entre paleontólogos de invertebrados. Los neandertales se convirtieron en una especie distinta del Homo sapiens, y recuperaron el nombre de Homo neanderthalensis, que había caído en desuso.
Los neandertales evolucionaron en Europa, por completo al margen de la evolución de la especie humana actual, y alcanzaron un grado muy apreciable de diferenciación morfológica. Conservaron algunas de las características del antepasado común, como la robustez y grosor de los huesos y la gran anchura del cuerpo. Pero acortaron los segmentos distales de las extremidades, es decir, el antebrazo y la tibia. En el cráneo mantuvieron la frente inclinada y la bóveda baja, pero expandieron el cerebro, ensanchando y alargando hacia atrás la caja craneana. También se modificó la forma de la cara, que se acuñó. A éstos se unieron otros muchos rasgos, como el espacio retromolar y la fosa suprainíaca del occipital. En el hombre moderno se estrechó el cilindro corporal, se aligeraron los huesos y la caja craneal se volvió esférica como respuesta a la expansión cerebral. El mentón de la mandíbula, el acortamiento de la cara y su reducción general son otros rasgos a destacar.
El proceso de divergencia entre neandertales y humanos actuales se inició hace medio millón de años, aproximadamente, pero en la época de la Sima de los Huesos en Atapuerca (en torno a los 300.000 años), las diferencias entre las poblaciones europeas y africanas eran todavía muy pequeñas. Seguramente formaban aún una misma especie biológica, que muchos autores modernos denominan Homo heidelbergensis (recuperando otro nombre antiguo, que fue creado para la mandíbula de Mauer, en Heidelberg, de hace 500.000 años).
Si los neandertales y nuestros antepasados de aquel tiempo pertenecían a la misma especie, entonces, por definición, no se podían cruzar, no podían intercambiar genes. ¿Cómo saberlo? Hay que empezar por decir que la diferencia morfológica (aunque sólo conocemos la del esqueleto) entre neandertales y humanos modernos es grande, y supera la observada entre especies próximas de primates y de mamíferos en general. Pero no es ésta prueba suficiente de aislamiento genético, ya que las barreras al intercambio de genes no se levantan simplemente como consecuencia de la acumulación de cambios morfológicos. A veces surgen especies nuevas sin que se produzca mucho cambio morfológico. Hay un acuerdo bastante general en que la aparición de barreras para el cruzamiento (sean de incompatibilidad en el comportamiento sexual, en los órganos y en la fisiología de la reproducción, o en los cromosomas) se da en poblaciones que ya estaban previamente aisladas geográficamente. O dicho con otras palabras, la poblaciones que no pueden intercambiar genes a causa de barreras físicas, terminan por no poder intercambiar genes por barreras biológicas.
Los neandertales evolucionaron durante cientos de miles de años en Europa, mientras nuestros antepasados lo hacía fuera del continente (seguramente en África, aunque el papel del Asia central y occidental aún queda por establecer). Es decir, que se dieron las condiciones de aislamiento geográfico necesarias (aunque no siempre suficientes) para que se produjera el aislamiento genético. Si se dio o no ya es otra cosa, y es lo que está por ver. Pero para ello se necesitan fósiles.
Los primeros fósiles de humanos modernos en Europa son posiblemente los de Mlade... [sic], en Moravia, República Checa. Su edad podría rondar los 32.000 años y se asocian a una industria Auriñaciense. Su morfología es rotundamente de Homo sapiens, y en nuestra opinión no hay nada en ellos que señale una relación con los neandertales, bien sea de descendencia directa, o bien de cruce entre ellos y los humanos modernos venidos de otra parte. Lo mismo puede decirse de otros fósiles seguramente algo más modernos, como los del yacimiento de Cro-Magnon. Ahora bien, estos fósiles modernos son posteriores en varios millares de años a los últimos neandertales de su región. Sería preferible estudiar a los humanos modernos que convivieron con los neandertales, o a los inmediatamente posteriores a su desaparición.
Como hemos comentado, la transición entre la industria Musteriense y las del Paleolítico Superior se produjo en los yacimientos del levante y sur de la Península bastante tiempo después que en el norte. Por eso los primeros humanos modernos que llegaron a la región no tendrán seguramente más de 30.000 años.El arqueólogo João Zilhão, el paleoantropólogo Erir Trinkaus y otros colegas anunciaron el descubrimiento de un resto infantil (de unos 4 años) en Lagar Velho (Portugal) que asignan al tecnocomplejo Gravetiense y datan en unos 24.500 años. Aunque se trata de un Homo sapiens, este individuo habría conservado las proporciones corporales (con acortamiento de la tibia) de un supuesto antepasado neandertal (aunque de todos modos anterior en muchas generaciones). Hay abundantes restos humanos en Moravia, Italia y otros yacimientos de Europa asociados a ese mismo tecnocomplejo (Gravetiense), pero no presentan proporciones corporales neandertales. Sin embargo, la reciente extinción de los neandertales en Portugal sitúa al niño de Lagar Velho más cerca de la época en la que los dos tipos humanos coincidieron y tal vez las aportaciones genéticas de los neandertales no se hubieran todavía perdido. Eso al menos es lo que afirman los citados autores, pero en nuestra opinión no se han presentado argumentos morfológicos suficientes a partir del niño de Lagar Velho para probar tal hipótesis. Otros autores, como Ian Tattersall y Jeffrey H. Schwartz, tampoco la admiten.
En el País Valenciano hay un importante resto de esa misma época, incluso algo anterior. Se trata del occipital de Malladetes, también encontrado en un contexto gravetiense. Tenemos pues la oportunidad de poner a prueba la tesis de la hibridación entre neandertales y humanos modernos. El occipital está muy completo, lo que es una buena noticia. La mala es que es infantil, por lo que muchas de las estructuras que diferencian a humanos modernos de neandertales aún no habrían tenido tiempo de desarrollarse. Hay sin embargo una que aparece muy pronto: se trata de la fosa suprainíaca, que se detecta hasta en niños muy pequeños entre los neandertales, y falta por completo en los humanos modernos, sean niños o adultos. Sin embargo falta en Malladetes. La consecuencia es que este occipital perteneció sin duda a un humano moderno, y no a un neandertal. Otra cosa muy diferente es imaginarse como sería la morfología occipital de un niño moderno Gravetiense perteneciente a una población que varios miles de años antes (entre dos mil y cinco mil) recibió una aportación (seguramente pequeña) de genes neandertales. Se trata pues de un ejercicio de ciencia ficción, en el que es mejor no entrar. La hipótesis de la asimilación de genes neandertales no se puede considerar probada, aunque todavía es escaso el registro de los primeros momentos de la ocupación humana moderna del sur de la Península Ibérica y futuros hallazgos pueden producir sorpresas.
2.5.2. Homo sapiens idaltu.
El Homo sapiens idaltu es una subespecie de Homo sapiens descrita a partir de especímenes fósiles encontrados en las proximidades de la localidad etíope de Herto en el año 1997 por el equipo de paleoantropólogos dirigido por Tim White. Sin embargo, el nuevo descubrimiento no se hizo público hasta el año 2003.
Dado el lugar en el cual se hizo el hallazgo de los restos (tres cráneos) inicialmente se le llamó Hombre de Herto. Dichos fósiles han sido datados con una antigüedad de unos 158.000 años, es decir, del Pleistoceno Medio.
El descubrimiento ha suscitado alguna polémica entre los investigadores. Varios consideran al Homo sapiens idaltu como una subespecie extinta del Homo sapiens y es por esto que se usa la denominación taxonómica trinomial. Otros, en cambio, consideran que Homo sapiens idaltu es sólo un ser humano de la actual especie que mantiene ligeros rasgos morfológicos arcaicos en relación a otros fósiles que son indiscutiblemente de Homo sapiens.
Si es correcta la tesitura según la cual el Homo sapiens idaltu es una subespecie de la nuestra, habrá que rescatar la antigua denominación de Homo sapiens sapiens para el hombre actual. Este terminología se usaba cuando el Hombre de Neandertal era considerado una subespecie nuestra (Homo sapiens neanderthalensis); pero desde que se evidenció que los neandertales y los humanos modernos representan especies biológicas distintas, se han venido usando los términos Homo sapiens y Homo neanderthalensis.


